

MediMer: a versatile do-it-yourself peptide-receptive MHC class I multimer platform for tumor neoantigen-specific T cell detection
- PMID: 38239352
- PMCID: PMC10794645
- DOI: 10.3389/fimmu.2023.1294565
MediMer: a versatile do-it-yourself peptide-receptive MHC class I multimer platform for tumor neoantigen-specific T cell detection
Abstract
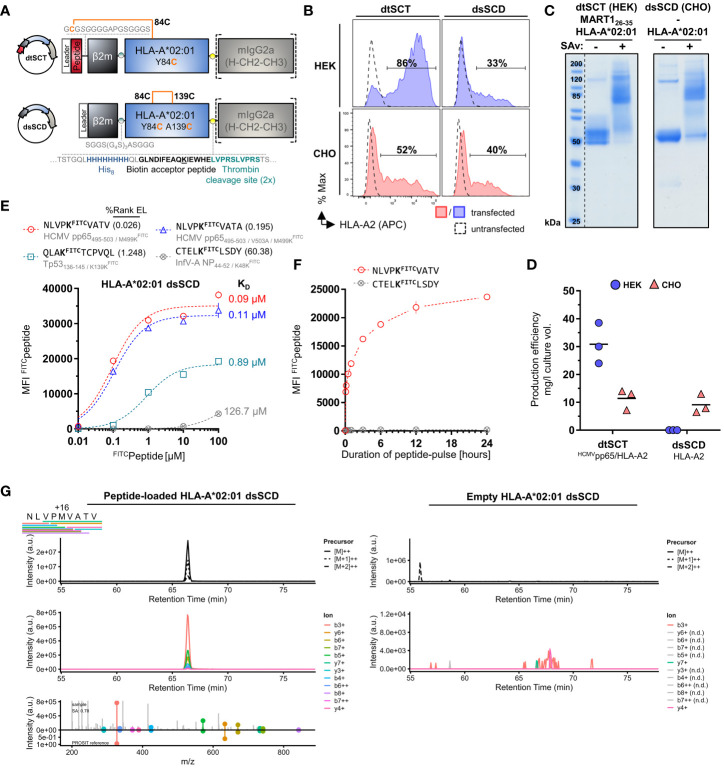
Peptide-loaded MHC class I (pMHC-I) multimers have revolutionized our capabilities to monitor disease-associated T cell responses with high sensitivity and specificity. To improve the discovery of T cell receptors (TCR) targeting neoantigens of individual tumor patients with recombinant MHC molecules, we developed a peptide-loadable MHC class I platform termed MediMer. MediMers are based on soluble disulfide-stabilized β2-microglobulin/heavy chain ectodomain single-chain dimers (dsSCD) that can be easily produced in large quantities in eukaryotic cells and tailored to individual patients' HLA allotypes with only little hands-on time. Upon transient expression in CHO-S cells together with ER-targeted BirA biotin ligase, biotinylated dsSCD are purified from the cell supernatant and are ready to use. We show that CHO-produced dsSCD are free of endogenous peptide ligands. Empty dsSCD from more than 30 different HLA-A,B,C allotypes, that were produced and validated so far, can be loaded with synthetic peptides matching the known binding criteria of the respective allotypes, and stored at low temperature without loss of binding activity. We demonstrate the usability of peptide-loaded dsSCD multimers for the detection of human antigen-specific T cells with comparable sensitivities as multimers generated with peptide-tethered β2m-HLA heavy chain single-chain trimers (SCT) and wild-type peptide-MHC-I complexes prior formed in small-scale refolding reactions. Using allotype-specific, fluorophore-labeled competitor peptides, we present a novel dsSCD-based peptide binding assay capable of interrogating large libraries of in silico predicted neoepitope peptides by flow cytometry in a high-throughput and rapid format. We discovered rare T cell populations with specificity for tumor neoepitopes and epitopes from shared tumor-associated antigens in peripheral blood of a melanoma patient including a so far unreported HLA-C*08:02-restricted NY-ESO-1-specific CD8+ T cell population. Two representative TCR of this T cell population, which could be of potential value for a broader spectrum of patients, were identified by dsSCD-guided single-cell sequencing and were validated by cognate pMHC-I multimer staining and functional responses to autologous peptide-pulsed antigen presenting cells. By deploying the technically accessible dsSCD MHC-I MediMer platform, we hope to significantly improve success rates for the discovery of personalized neoepitope-specific TCR in the future by being able to also cover rare HLA allotypes.
Keywords: T cell receptor discovery; T cells; neoepitope screening; peptide-MHC class I multimer; personalized medicine; tumor immunotherapy; tumor neoantigen.
Copyright © 2024 Meyer, Parpoulas, Barthélémy, Becker, Charoentong, Lyu, Börsig, Bulbuc, Tessmer, Weinacht, Ibberson, Schmidt, Pipkorn, Eichmüller, Steinberger, Lindner, Poschke, Platten, Fröhling, Riemer, Hassel, Roberti, Jäger, Zörnig and Momburg.
Conflict of interest statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest. The author(s) declared that they were an editorial board member of Frontiers, at the time of submission. This had no impact on the peer review process and the final decision.
Figures






References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials