

MCJ: A mitochondrial target for cardiac intervention in pulmonary hypertension
- PMID: 38241373
- PMCID: PMC10798563
- DOI: 10.1126/sciadv.adk6524
MCJ: A mitochondrial target for cardiac intervention in pulmonary hypertension
Abstract
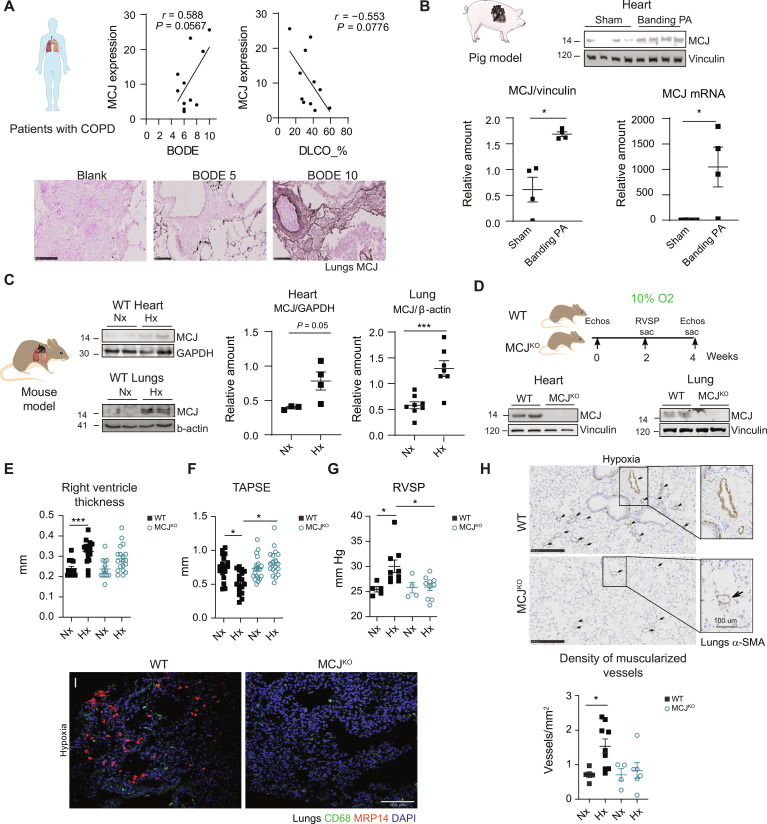
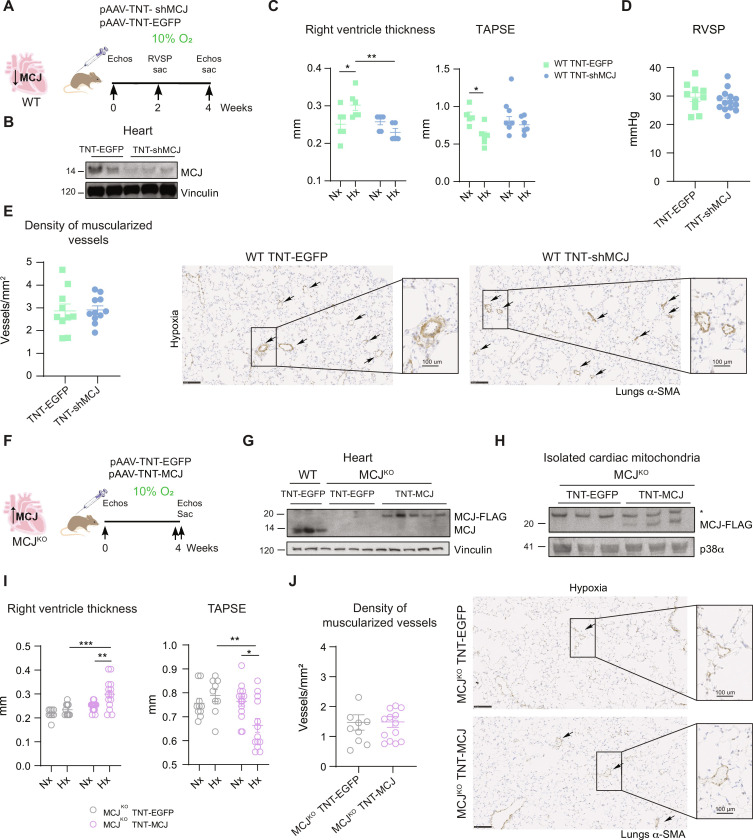
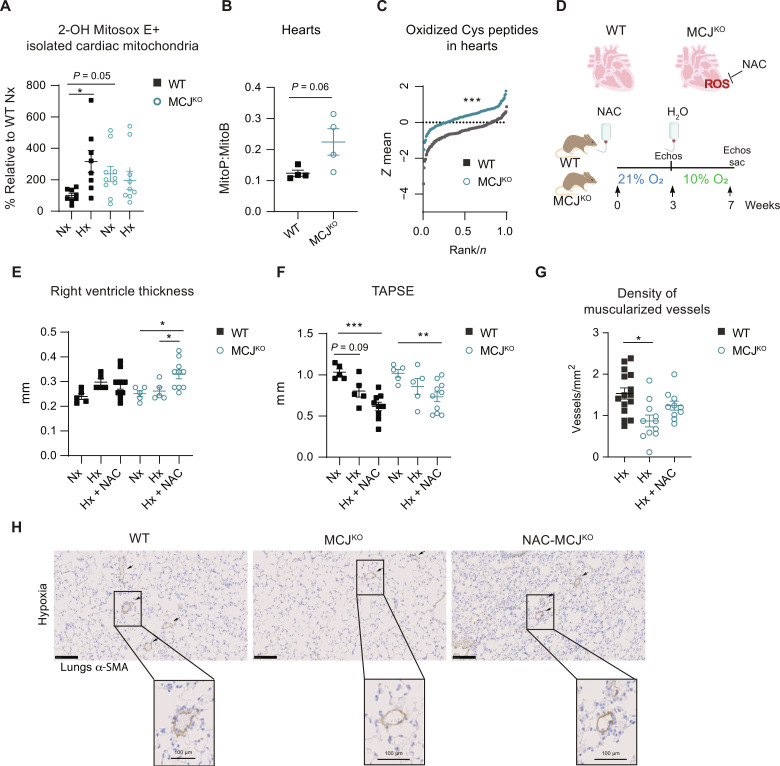
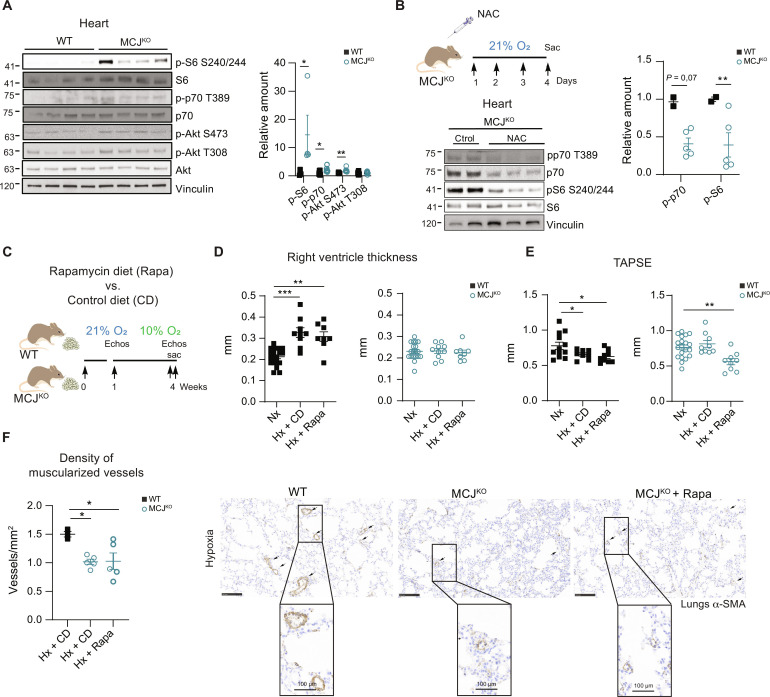
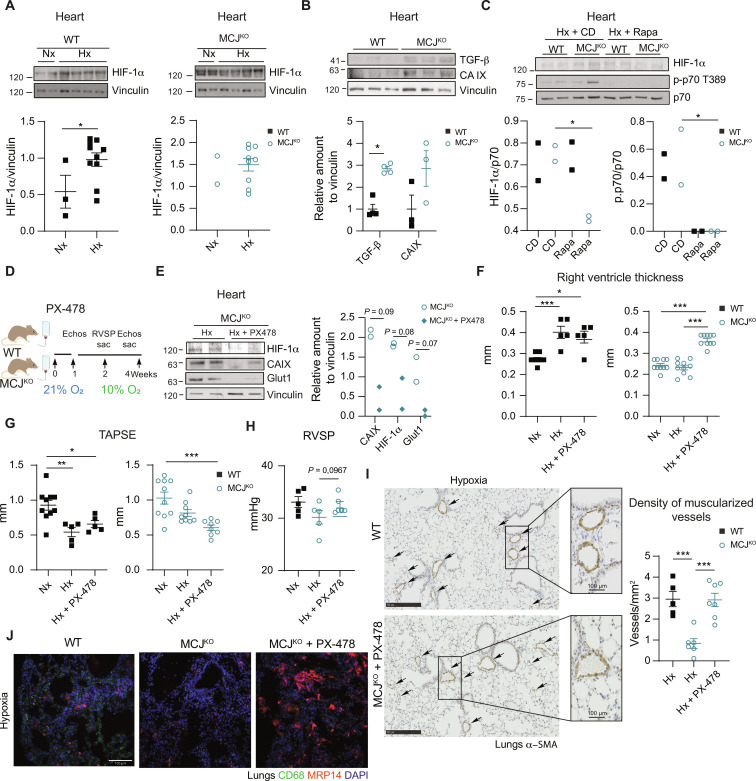
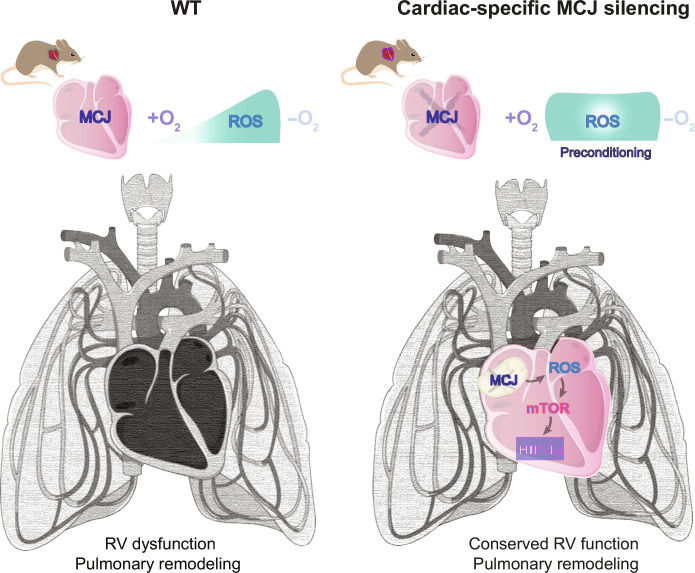
Pulmonary hypertension (PH) can affect both pulmonary arterial tree and cardiac function, often leading to right heart failure and death. Despite the urgency, the lack of understanding has limited the development of effective cardiac therapeutic strategies. Our research reveals that MCJ modulates mitochondrial response to chronic hypoxia. MCJ levels elevate under hypoxic conditions, as in lungs of patients affected by COPD, mice exposed to hypoxia, and myocardium from pigs subjected to right ventricular (RV) overload. The absence of MCJ preserves RV function, safeguarding against both cardiac and lung remodeling induced by chronic hypoxia. Cardiac-specific silencing is enough to protect against cardiac dysfunction despite the adverse pulmonary remodeling. Mechanistically, the absence of MCJ triggers a protective preconditioning state mediated by the ROS/mTOR/HIF-1α axis. As a result, it preserves RV systolic function following hypoxia exposure. These discoveries provide a potential avenue to alleviate chronic hypoxia-induced PH, highlighting MCJ as a promising target against this condition.
Figures
References
-
- Arias-Stella J., Saldana M., The terminal portion of the pulmonary arterial tree in people native to high altitudes. Circulation 28, 915–925 (1963). - PubMed
-
- Zhao L., Chronic hypoxia-induced pulmonary hypertension in rat: The best animal model for studying pulmonary vasoconstriction and vascular medial hypertrophy. Drug Discov. Today Dis. Model. 7, 83–88 (2010).
-
- Humbert M., Morrell N. W., Archer S. L., Stenmark K. R., MacLean M. R., Lang I. M., Christman B. W., Weir E. K., Eickelberg O., Voelkel N. F., Rabinovitch M., Cellular and molecular pathobiology of pulmonary arterial hypertension. J. Am. Coll. Cardiol. 43, 13S–24S (2004). - PubMed
-
- Humbert M., Guignabert C., Bonnet S., Dorfmüller P., Klinger J. R., Nicolls M. R., Olschewski A. J., Pullamsetti S. S., Schermuly R. T., Stenmark K. R., Rabinovitch M., Pathology and pathobiology of pulmonary hypertension: State of the art and research perspectives. Eur. Respir. J. 53, 1801887 (2019). - PMC - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
