

Transcription regulation by long non-coding RNAs: mechanisms and disease relevance
- PMID: 38242953
- PMCID: PMC11045326
- DOI: 10.1038/s41580-023-00694-9
Transcription regulation by long non-coding RNAs: mechanisms and disease relevance
Abstract
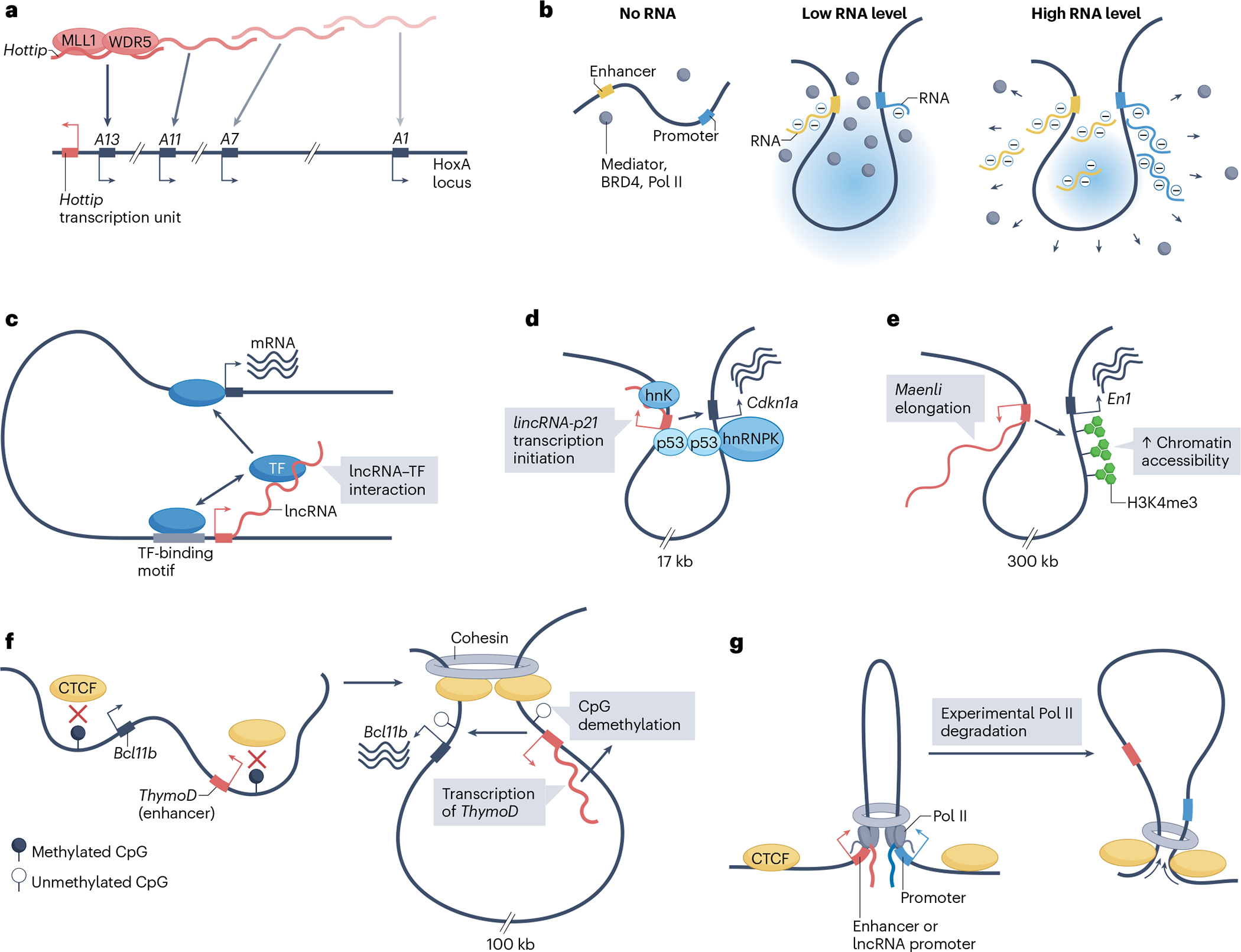
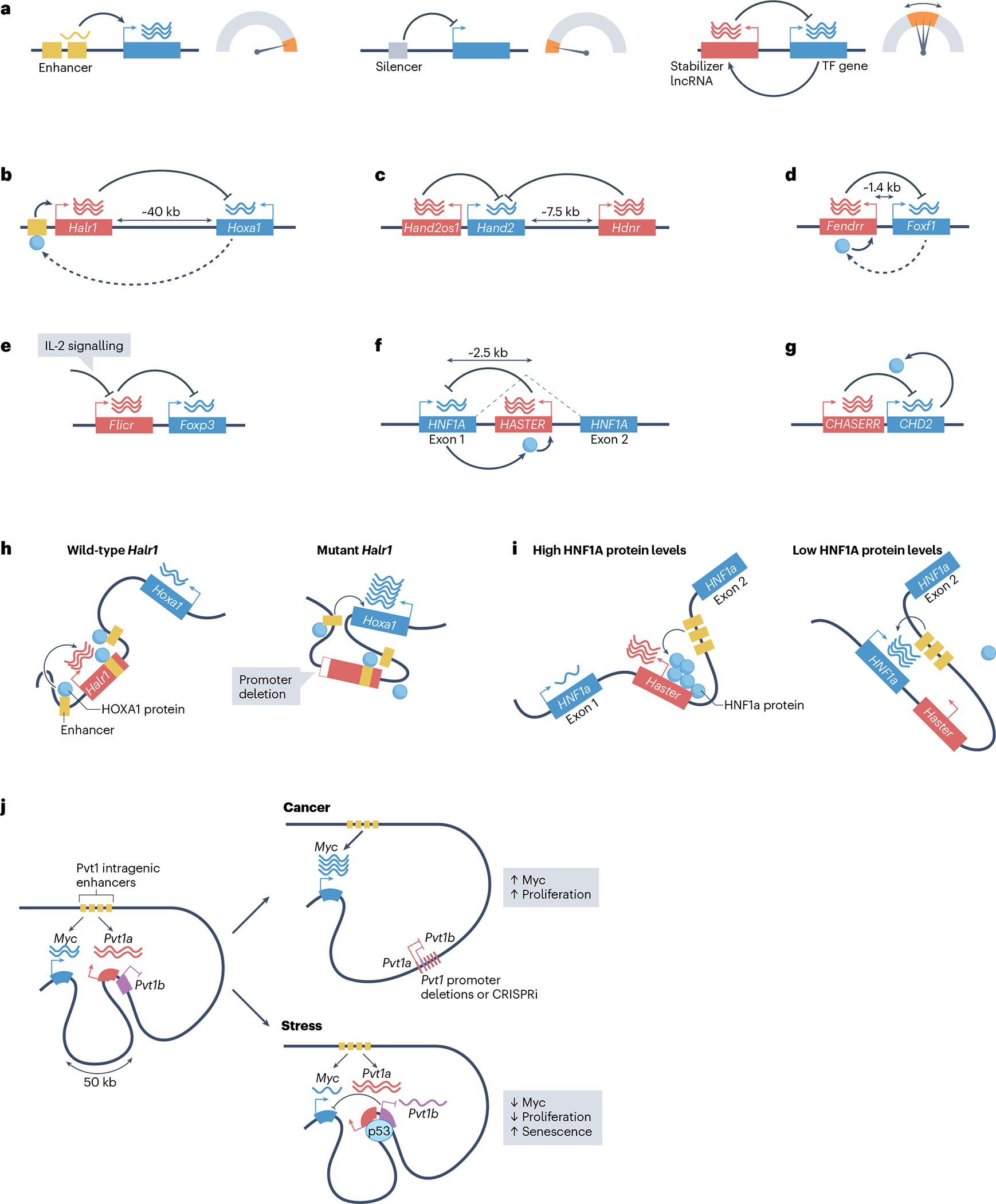
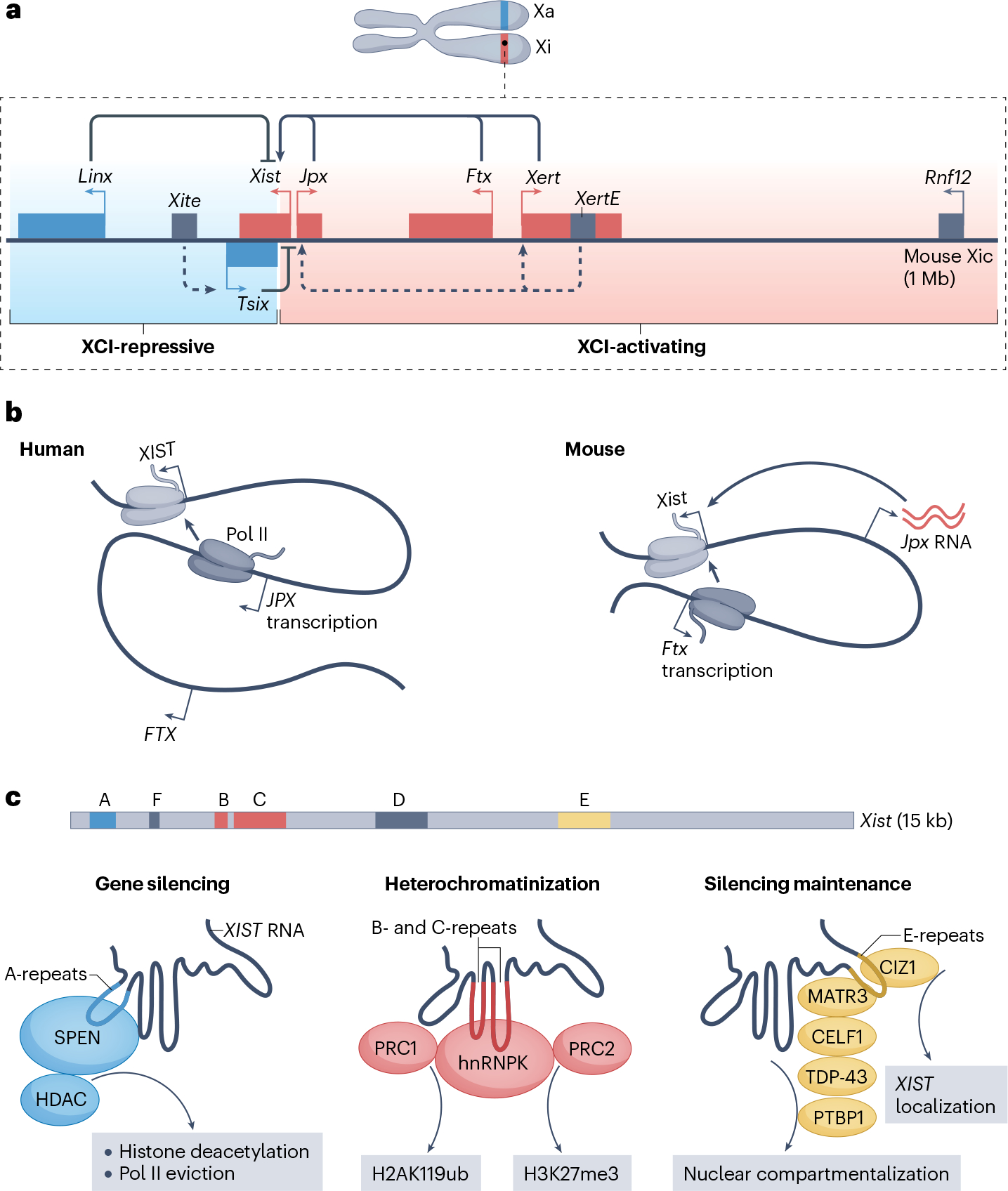
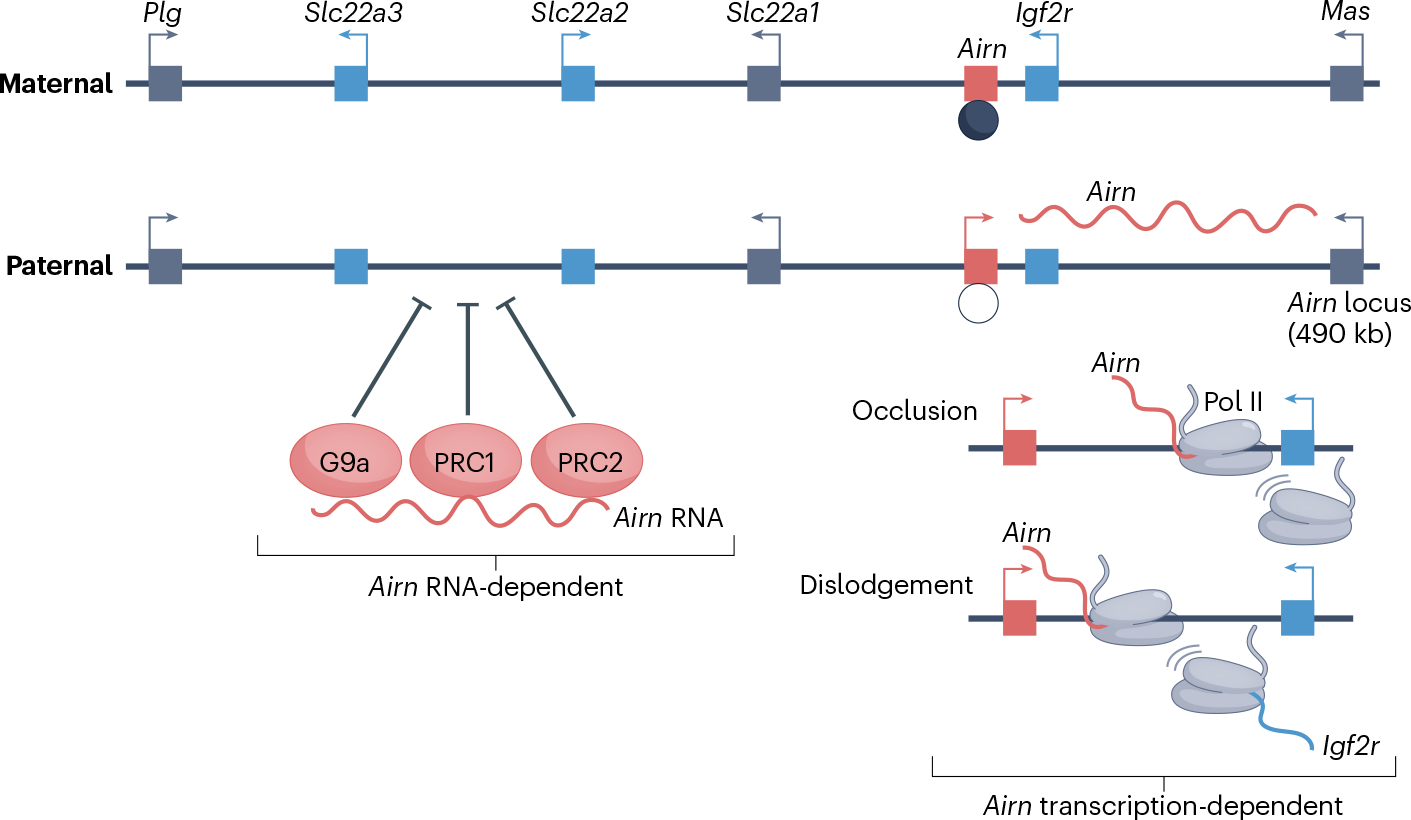
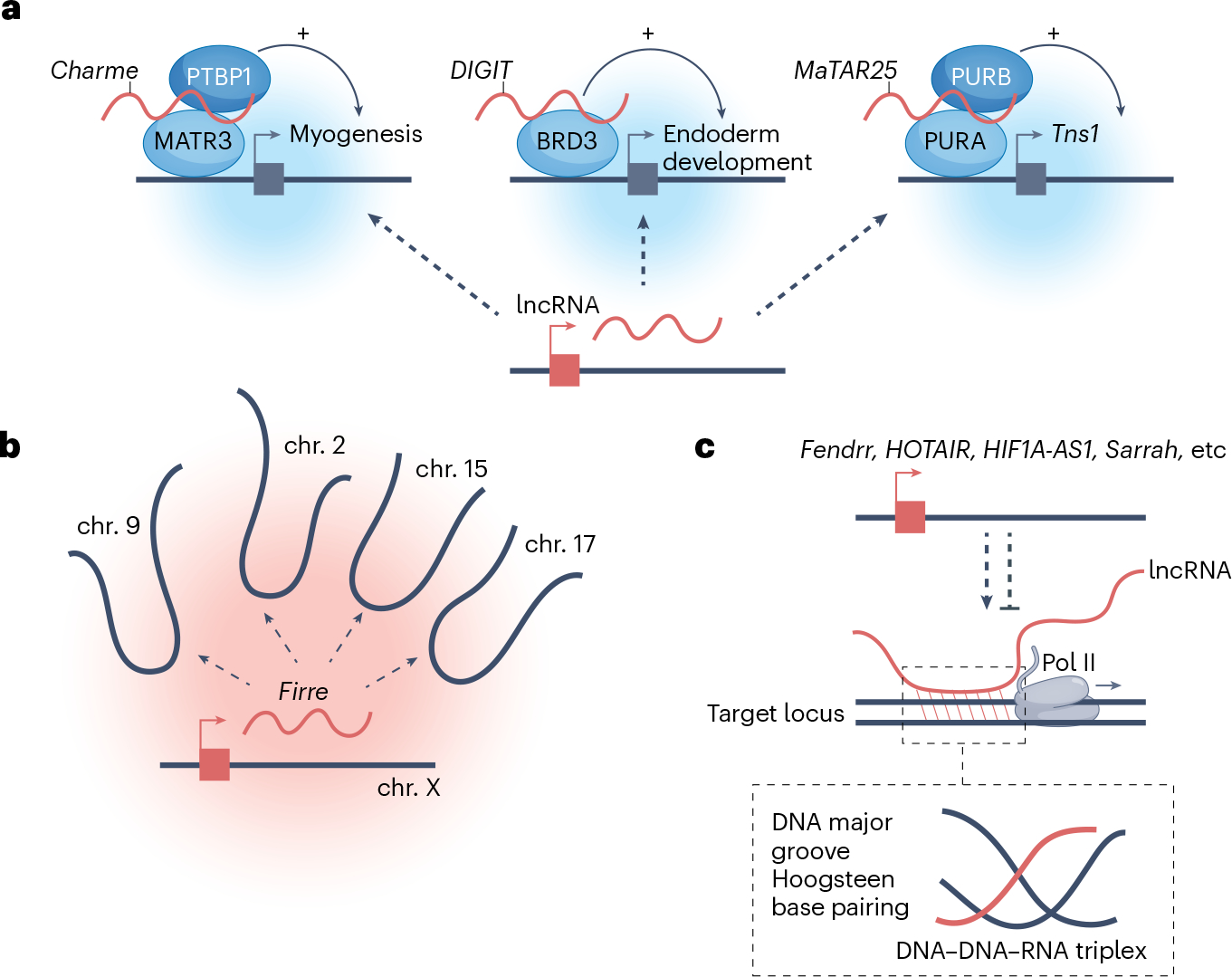
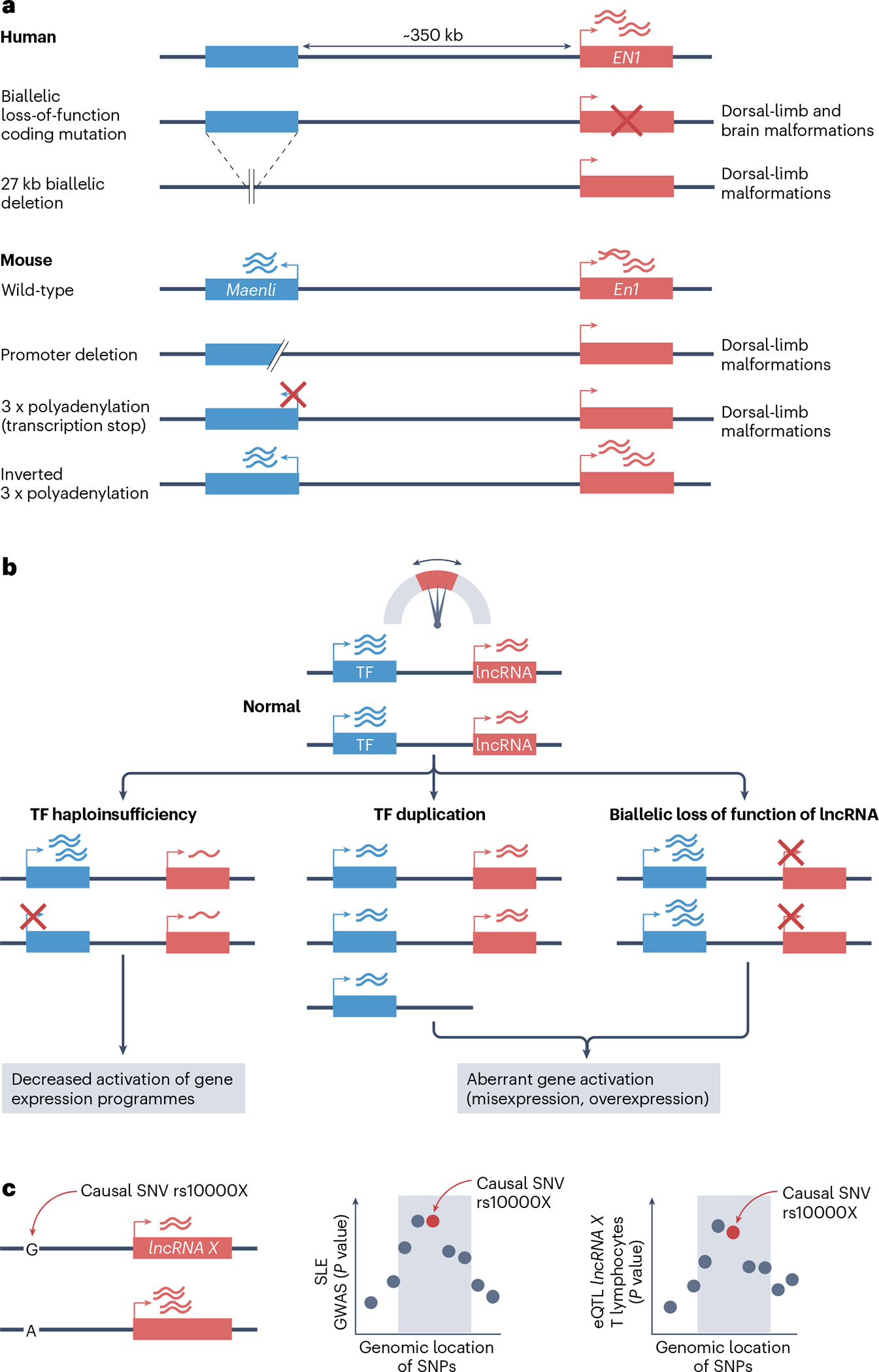
Long non-coding RNAs (lncRNAs) outnumber protein-coding transcripts, but their functions remain largely unknown. In this Review, we discuss the emerging roles of lncRNAs in the control of gene transcription. Some of the best characterized lncRNAs have essential transcription cis-regulatory functions that cannot be easily accomplished by DNA-interacting transcription factors, such as XIST, which controls X-chromosome inactivation, or imprinted lncRNAs that direct allele-specific repression. A growing number of lncRNA transcription units, including CHASERR, PVT1 and HASTER (also known as HNF1A-AS1) act as transcription-stabilizing elements that fine-tune the activity of dosage-sensitive genes that encode transcription factors. Genetic experiments have shown that defects in such transcription stabilizers often cause severe phenotypes. Other lncRNAs, such as lincRNA-p21 (also known as Trp53cor1) and Maenli (Gm29348) contribute to local activation of gene transcription, whereas distinct lncRNAs influence gene transcription in trans. We discuss findings of lncRNAs that elicit a function through either activation of their transcription, transcript elongation and processing or the lncRNA molecule itself. We also discuss emerging evidence of lncRNA involvement in human diseases, and their potential as therapeutic targets.
© 2024. Springer Nature Limited.
Figures
References
-
- Lipshitz HD, Peattie DA & Hogness DS Novel transcripts from the ultrabithorax domain of the bithorax complex. Genes. Dev. 1, 307–322 (1987). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
