

Extracellular lipidosomes containing lipid droplets and mitochondria are released during melanoma cell division
- PMID: 38243233
- PMCID: PMC10799373
- DOI: 10.1186/s12964-024-01471-7
Extracellular lipidosomes containing lipid droplets and mitochondria are released during melanoma cell division
Abstract
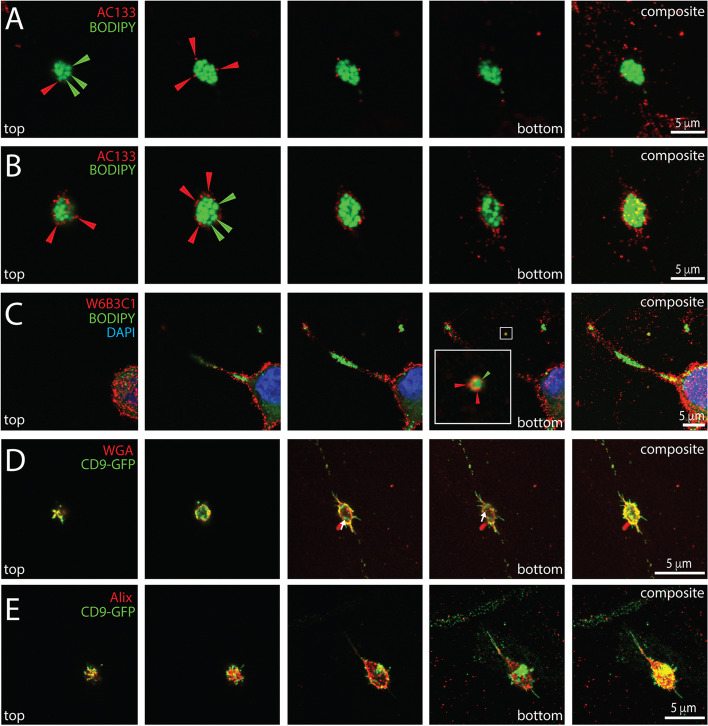
Background: The incidence of melanoma is increasing worldwide. Since metastatic melanoma is highly aggressive, it is important to decipher all the biological aspects of melanoma cells. In this context, we have previously shown that metastatic FEMX-I melanoma cells release small (< 150 nm) extracellular vesicles (EVs) known as exosomes and ectosomes containing the stem (and cancer stem) cell antigenic marker CD133. EVs play an important role in intercellular communication, which could have a micro-environmental impact on surrounding tissues.
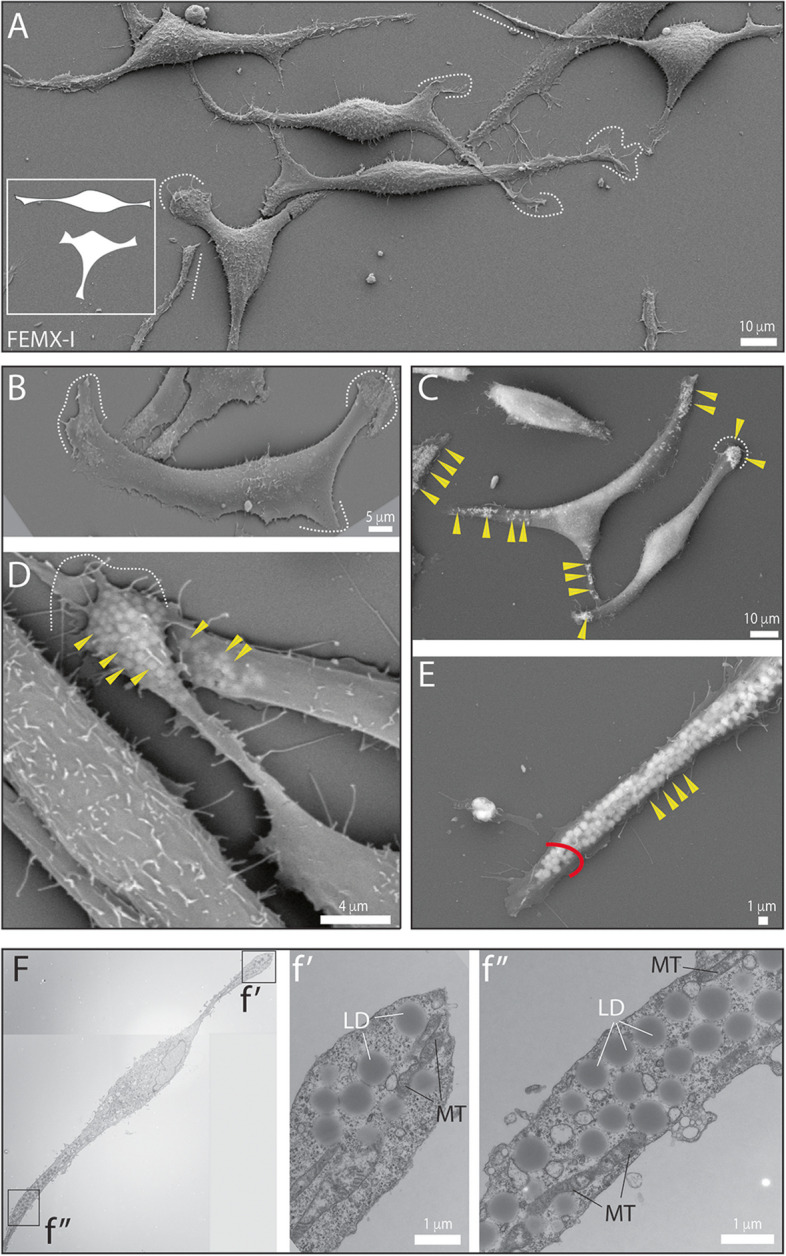
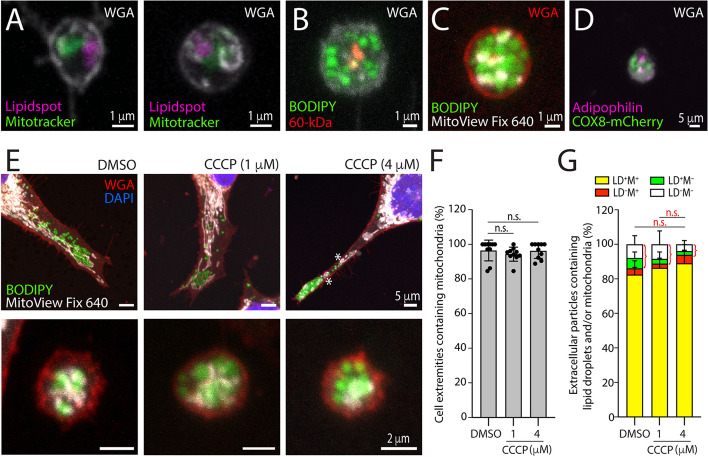
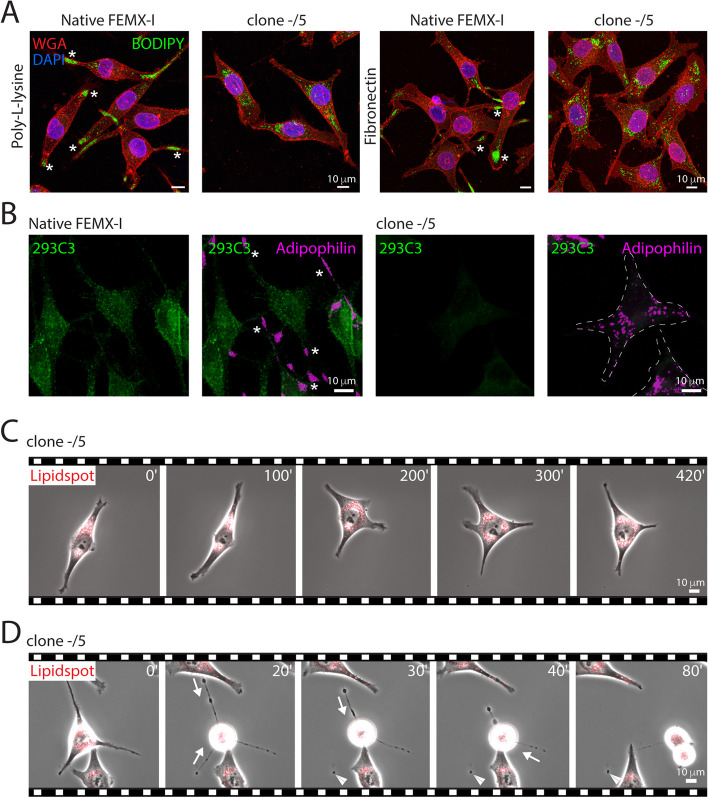
Results: We report here a new type of large CD133+ EVs released by FEMX-I cells. Their sizes range from 2 to 6 µm and they contain lipid droplets and mitochondria. Real-time video microscopy revealed that these EVs originate from the lipid droplet-enriched cell extremities that did not completely retract during the cell division process. Once released, they can be taken up by other cells. Silencing CD133 significantly affected the cellular distribution of lipid droplets, with a re-localization around the nuclear compartment. As a result, the formation of large EVs containing lipid droplets was severely compromised.
Conclusion: Given the biochemical effect of lipid droplets and mitochondria and/or their complexes on cell metabolism, the release and uptake of these new large CD133+ EVs from dividing aggressive melanoma cells can influence both donor and recipient cells, and therefore impact melanoma growth and dissemination.
Keywords: Cell division; Extracellular vesicle; Lipid droplet; Melanoma; Mitochondrion; Prominin-1.
© 2024. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures






References
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Research Materials
