

Calciprotein Particles Induce Cellular Compartment-Specific Proteome Alterations in Human Arterial Endothelial Cells
- PMID: 38248875
- PMCID: PMC10816121
- DOI: 10.3390/jcdd11010005
Calciprotein Particles Induce Cellular Compartment-Specific Proteome Alterations in Human Arterial Endothelial Cells
Abstract
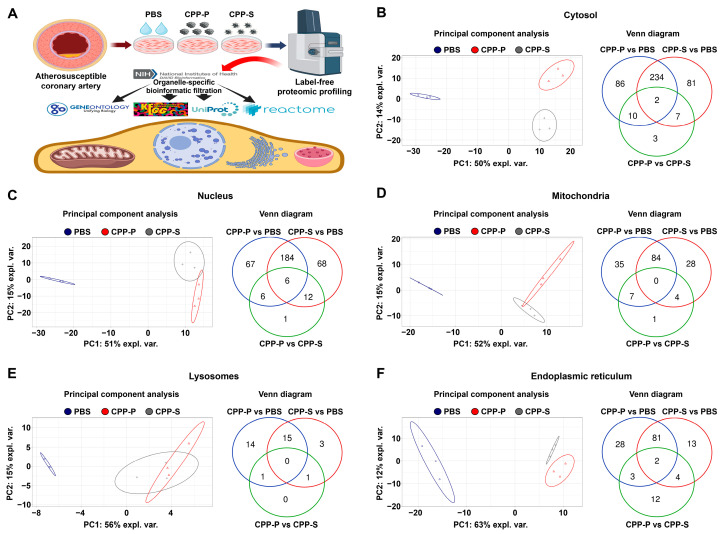
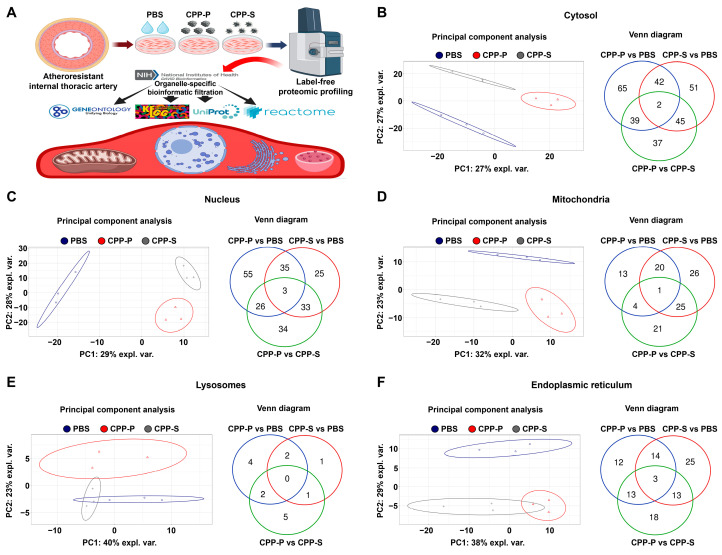
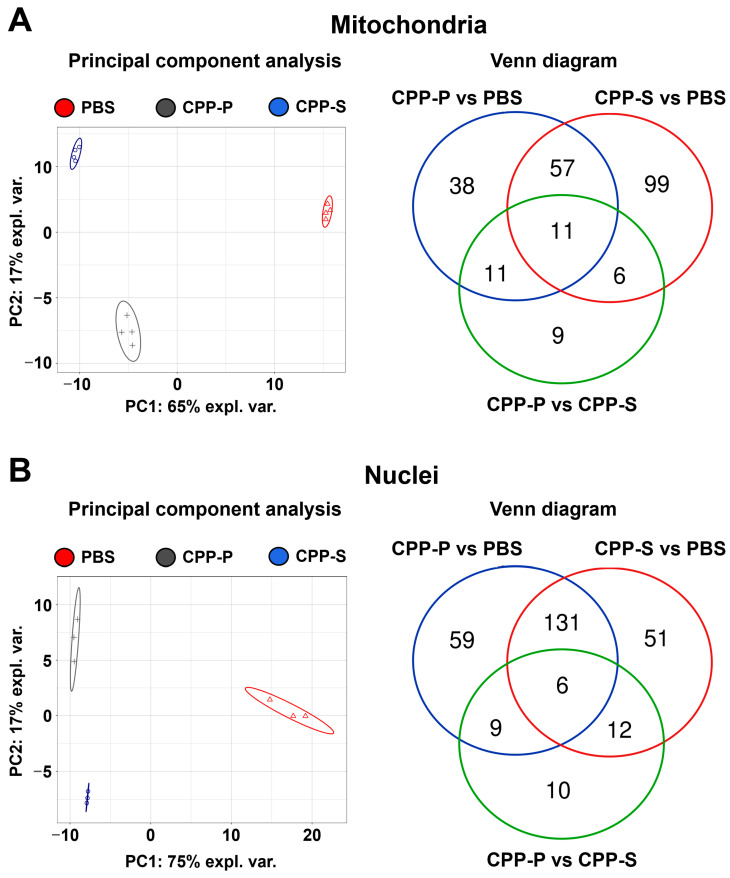
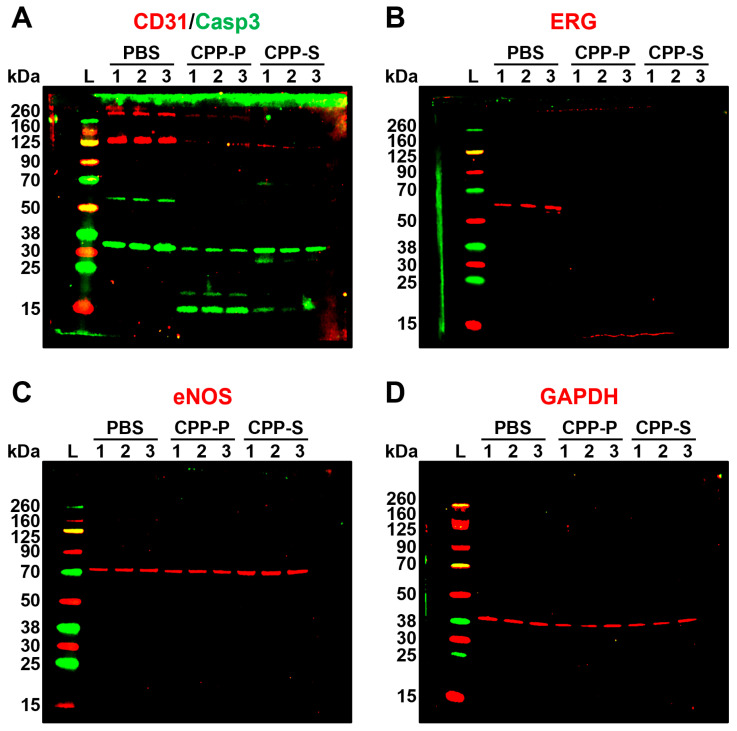
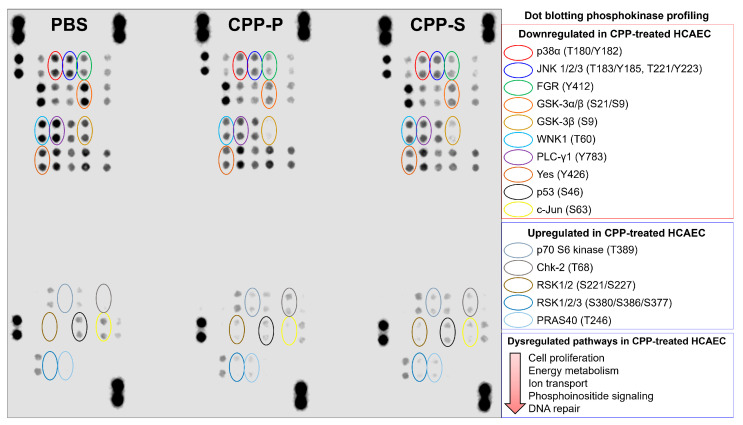
Calciprotein particles (CPPs) are indispensable scavengers of excessive Ca2+ and PO43- ions in blood, being internalised and recycled by liver and spleen macrophages, monocytes, and endothelial cells (ECs). Here, we performed a pathway enrichment analysis of cellular compartment-specific proteomes in primary human coronary artery ECs (HCAEC) and human internal thoracic artery ECs (HITAEC) treated with primary (amorphous) or secondary (crystalline) CPPs (CPP-P and CPPs, respectively). Exposure to CPP-P and CPP-S induced notable upregulation of: (1) cytokine- and chemokine-mediated signaling, Ca2+-dependent events, and apoptosis in cytosolic and nuclear proteomes; (2) H+ and Ca2+ transmembrane transport, generation of reactive oxygen species, mitochondrial outer membrane permeabilisation, and intrinsic apoptosis in the mitochondrial proteome; (3) oxidative, calcium, and endoplasmic reticulum (ER) stress, unfolded protein binding, and apoptosis in the ER proteome. In contrast, transcription, post-transcriptional regulation, translation, cell cycle, and cell-cell adhesion pathways were underrepresented in cytosol and nuclear compartments, whilst biosynthesis of amino acids, mitochondrial translation, fatty acid oxidation, pyruvate dehydrogenase activity, and energy generation were downregulated in the mitochondrial proteome of CPP-treated ECs. Differentially expressed organelle-specific pathways were coherent in HCAEC and HITAEC and between ECs treated with CPP-P or CPP-S. Proteomic analysis of mitochondrial and nuclear lysates from CPP-treated ECs confirmed bioinformatic filtration findings.
Keywords: calciprotein particles; cytosol; endoplasmic reticulum; endothelial cells; lysosomes; mineral stress; mitochondria; molecular signatures; nuclei; proteomic profiling.
Conflict of interest statement
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
Figures
Similar articles
-
Proteomic Profiling of Endothelial Cell Secretomes After Exposure to Calciprotein Particles Reveals Downregulation of Basement Membrane Assembly and Increased Release of Soluble CD59.Int J Mol Sci. 2024 Oct 23;25(21):11382. doi: 10.3390/ijms252111382. Int J Mol Sci. 2024. PMID: 39518935 Free PMC article.
-
Calciprotein particle-induced calcium overload triggers mitochondrial dysfunction in endothelial cells.J Physiol. 2025 Mar 9. doi: 10.1113/JP287656. Online ahead of print. J Physiol. 2025. PMID: 40059365
-
Calciprotein Particles Cause Physiologically Significant Pro-Inflammatory Response in Endothelial Cells and Systemic Circulation.Int J Mol Sci. 2022 Nov 29;23(23):14941. doi: 10.3390/ijms232314941. Int J Mol Sci. 2022. PMID: 36499266 Free PMC article.
-
Alterations in inter-organelle crosstalk and Ca2+ signaling through mitochondria during proteotoxic stresses.Mitochondrion. 2021 Mar;57:37-46. doi: 10.1016/j.mito.2020.12.003. Epub 2020 Dec 17. Mitochondrion. 2021. PMID: 33340711 Review.
-
Calciprotein particles: mineral behaving badly?Curr Opin Nephrol Hypertens. 2020 Jul;29(4):378-386. doi: 10.1097/MNH.0000000000000609. Curr Opin Nephrol Hypertens. 2020. PMID: 32404635 Review.
Cited by
-
The Bone-Vascular Axis: A Key Player in Chronic Kidney Disease Associated Vascular Calcification.Kidney Dis (Basel). 2024 Sep 6;10(6):545-557. doi: 10.1159/000541280. eCollection 2024 Dec. Kidney Dis (Basel). 2024. PMID: 39664335 Free PMC article. Review.
-
The Role of Cardio-Renal Inflammation in Deciding the Fate of the Arteriovenous Fistula in Haemodialysis Therapy.Cells. 2024 Oct 1;13(19):1637. doi: 10.3390/cells13191637. Cells. 2024. PMID: 39404400 Free PMC article. Review.
-
The formation and function of calciprotein particles.Pflugers Arch. 2025 Jun;477(6):753-772. doi: 10.1007/s00424-025-03083-7. Epub 2025 Apr 23. Pflugers Arch. 2025. PMID: 40266378 Free PMC article. Review.
-
Proteomic Profiling of Endothelial Cell Secretomes After Exposure to Calciprotein Particles Reveals Downregulation of Basement Membrane Assembly and Increased Release of Soluble CD59.Int J Mol Sci. 2024 Oct 23;25(21):11382. doi: 10.3390/ijms252111382. Int J Mol Sci. 2024. PMID: 39518935 Free PMC article.
References
-
- Heiss A., DuChesne A., Denecke B., Grötzinger J., Yamamoto K., Renné T., Jahnen-Dechent W. Structural basis of calcification inhibition by alpha 2-HS glycoprotein/fetuin-A. Formation of colloidal calciprotein particles. J. Biol. Chem. 2003;278:13333–13341. doi: 10.1074/jbc.M210868200. - DOI - PubMed
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
