

This is a preprint.
Reprogramming neuroblastoma by diet-enhanced polyamine depletion
- PMID: 38260457
- PMCID: PMC10802427
- DOI: 10.1101/2024.01.07.573662
Reprogramming neuroblastoma by diet-enhanced polyamine depletion
Update in
-
Reprogramming neuroblastoma by diet-enhanced polyamine depletion.Nature. 2025 Oct;646(8085):707-715. doi: 10.1038/s41586-025-09564-0. Epub 2025 Sep 24. Nature. 2025. PMID: 40993392 Free PMC article.
Abstract
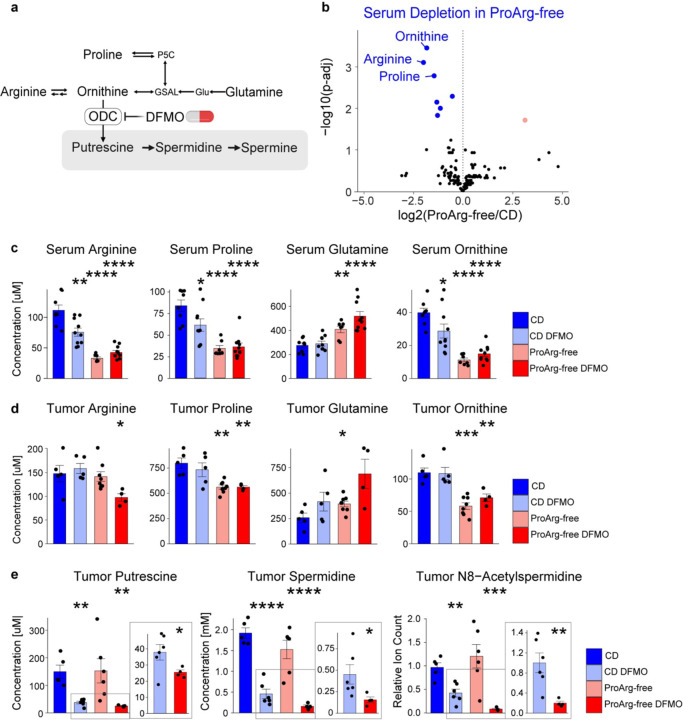
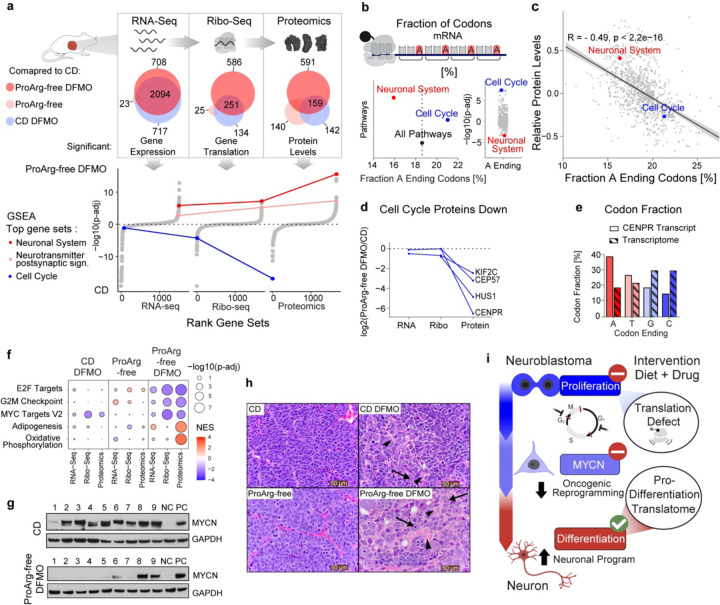
Neuroblastoma is a highly lethal childhood tumor derived from differentiation-arrested neural crest cells1,2. Like all cancers, its growth is fueled by metabolites obtained from either circulation or local biosynthesis3,4. Neuroblastomas depend on local polyamine biosynthesis, with the inhibitor difluoromethylornithine showing clinical activity5. Here we show that such inhibition can be augmented by dietary restriction of upstream amino acid substrates, leading to disruption of oncogenic protein translation, tumor differentiation, and profound survival gains in the TH-MYCN mouse model. Specifically, an arginine/proline-free diet decreases the polyamine precursor ornithine and augments tumor polyamine depletion by difluoromethylornithine. This polyamine depletion causes ribosome stalling, unexpectedly specifically at adenosine-ending codons. Such codons are selectively enriched in cell cycle genes and low in neuronal differentiation genes. Thus, impaired translation of these codons, induced by the diet-drug combination, favors a pro-differentiation proteome. These results suggest that the genes of specific cellular programs have evolved hallmark codon usage preferences that enable coherent translational rewiring in response to metabolic stresses, and that this process can be targeted to activate differentiation of pediatric cancers.
Keywords: Protein translation; amino acid restriction; arginine; diet; difluoromethylornithine; glutamine; ornithine; ornithine aminotransferase; polyamines; proline.
Conflict of interest statement
Competing interests J.D.R. is a member of the Rutgers Cancer Institute of New Jersey and the University of Pennsylvania Diabetes Research Center; a co-founder and stockholder in Empress Therapeutics and Serien Therapeutics; and an advisor and stockholder in Agios Pharmaceuticals, Bantam Pharmaceuticals, Colorado Research Partners, Rafael Pharmaceuticals, Barer Institute, and L.E.A.F. Pharmaceuticals. University of Zürich has filed a provisional patent on combining difluormethylornithine with amino acid manipulations for therapeutic use.
Figures



References
-
- Matthay K. K. et al. Neuroblastoma. Nat. Rev. Dis. Primer 2, 16078 (2016). - PubMed
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases