

This is a preprint.
SARS-CoV-2 papain-like protease activates nociceptors to drive sneeze and pain
- PMID: 38260476
- PMCID: PMC10802627
- DOI: 10.1101/2024.01.10.575114
SARS-CoV-2 papain-like protease activates nociceptors to drive sneeze and pain
Abstract
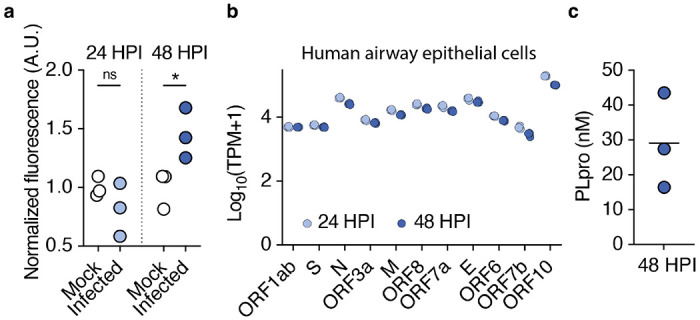
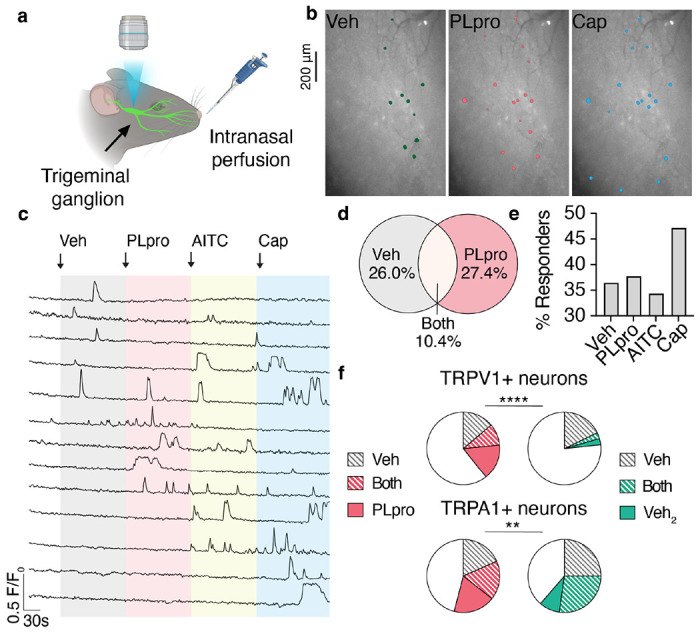
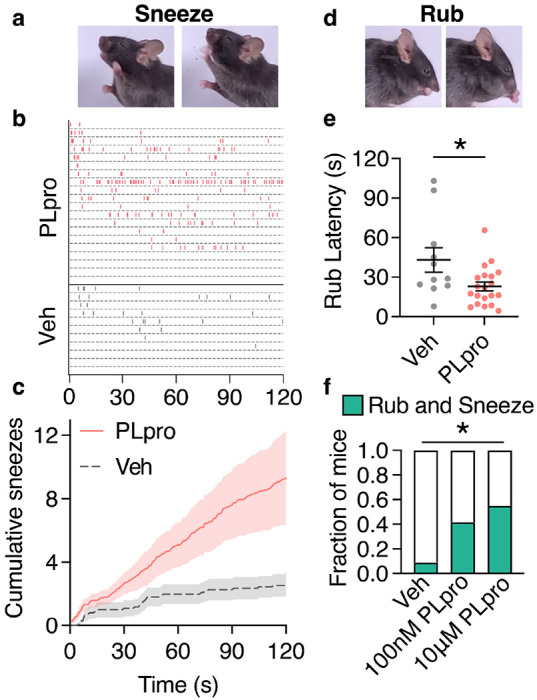
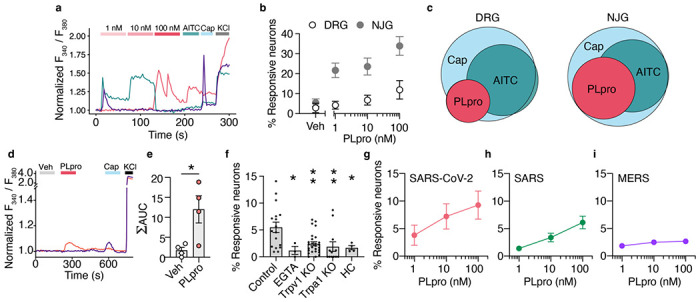
SARS-CoV-2, the virus responsible for COVID-19, triggers symptoms such as sneezing, aches and pain.1 These symptoms are mediated by a subset of sensory neurons, known as nociceptors, that detect noxious stimuli, densely innervate the airway epithelium, and interact with airway resident epithelial and immune cells.2-6 However, the mechanisms by which viral infection activates these neurons to trigger pain and airway reflexes are unknown. Here, we show that the coronavirus papain-like protease (PLpro) directly activates airway-innervating trigeminal and vagal nociceptors in mice and human iPSC-derived nociceptors. PLpro elicits sneezing and acute pain in mice and triggers the release of neuropeptide calcitonin gene-related peptide (CGRP) from airway afferents. We find that PLpro-induced sneeze and pain requires the host TRPA1 ion channel that has been previously demonstrated to mediate pain, cough, and airway inflammation.7-9 Our findings are the first demonstration of a viral product that directly activates sensory neurons to trigger pain and airway reflexes and highlight a new role for PLpro and nociceptors in COVID-19.
Figures
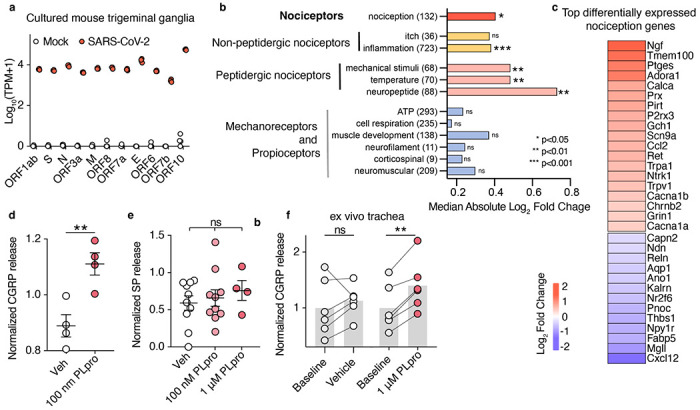
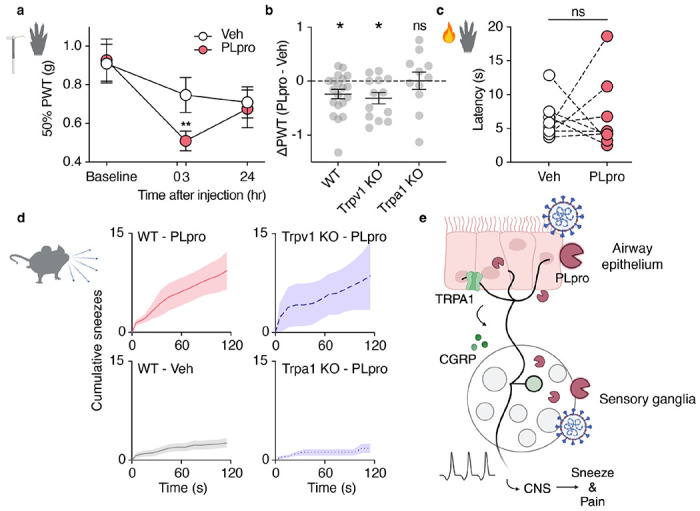
References
-
- Finger T. E., St Jeor V. L., Kinnamon J. C. & Silver W. L. Ultrastructure of substance P- and CGRP-immunoreactive nerve fibers in the nasal epithelium of rodents. J. Comp. Neurol. 294, 293–305 (1990). - PubMed
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous