

Ribosomal protein L24 mediates mammalian microRNA processing in an evolutionarily conserved manner
- PMID: 38261097
- PMCID: PMC10805976
- DOI: 10.1007/s00018-023-05088-w
Ribosomal protein L24 mediates mammalian microRNA processing in an evolutionarily conserved manner
Abstract
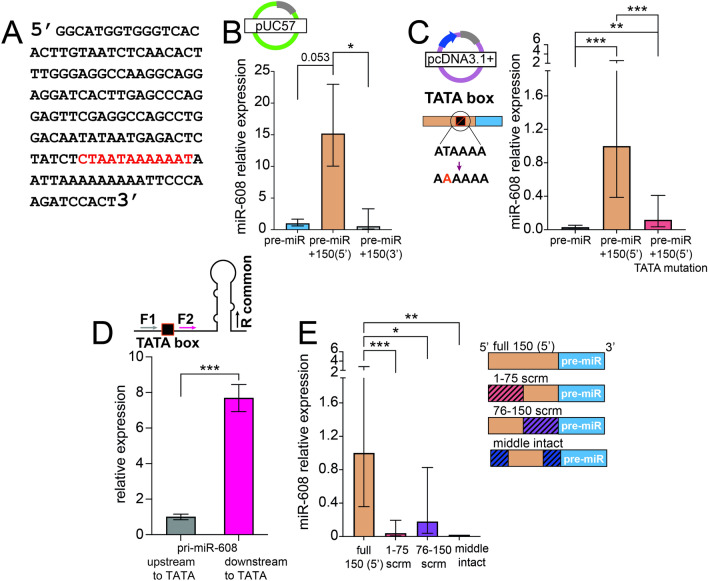
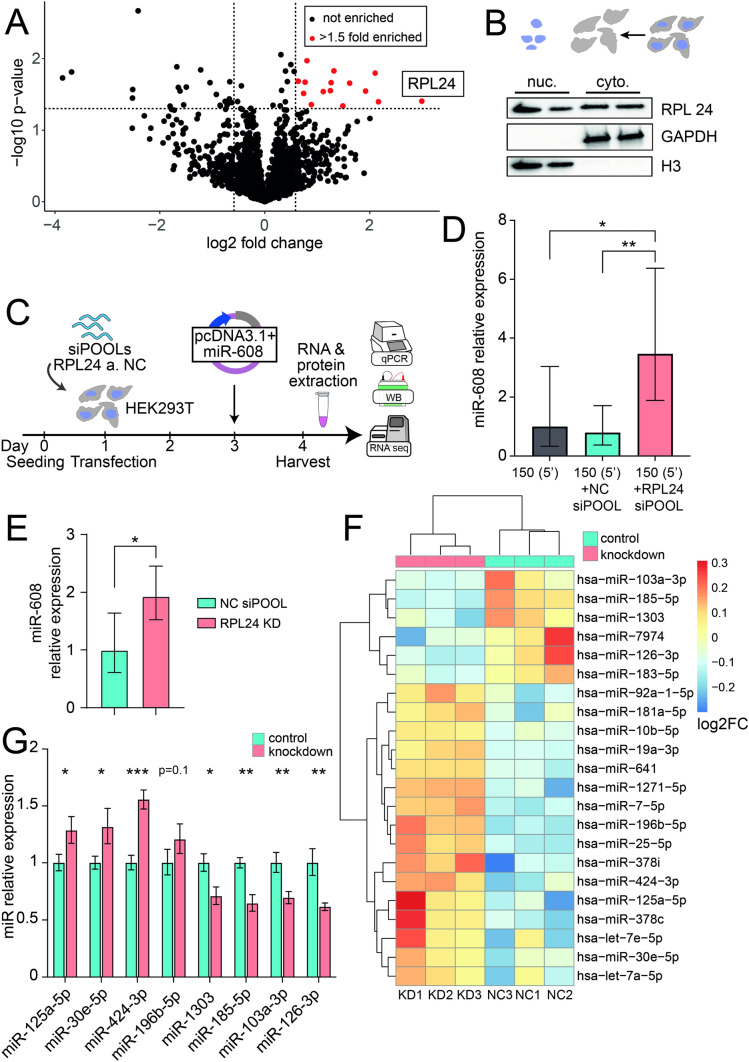
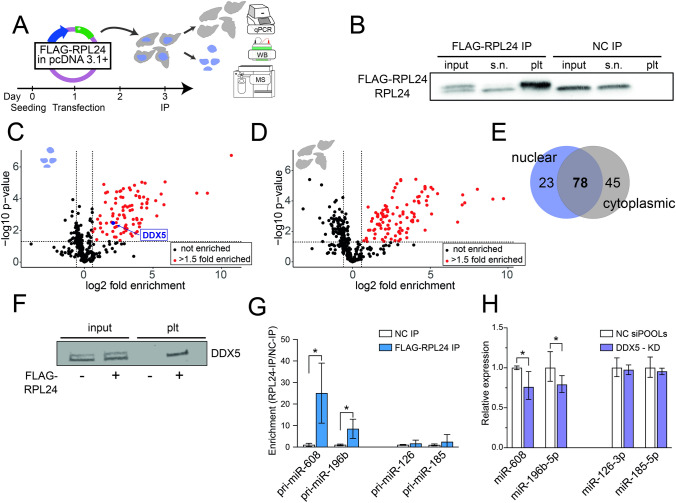
To investigate the mechanism(s) underlying the expression of primate-specific microRNAs (miRs), we sought DNA regulatory elements and proteins mediating expression of the primate-specific hsa-miR-608 (miR-608), which is located in the SEMA4G gene and facilitates the cholinergic blockade of inflammation by targeting acetylcholinesterase mRNA. 'Humanized' mice carrying pre-miR-608 flanked by 250 bases of endogenous sequences inserted into the murine Sema4g gene successfully expressed miR-608. Moreover, by flanking miR-608 by shortened fragments of its human genome region we identified an active independent promoter within the 150 nucleotides 5' to pre-miR-608, which elevated mature miR-608 levels by 100-fold in transfected mouse- and human-originated cells. This highlighted a regulatory role of the 5' flank as enabling miR-608 expression. Moreover, pull-down of the 150-base 5' sequence revealed its interaction with ribosomal protein L24 (RPL24), implicating an additional mechanism controlling miR-608 levels. Furthermore, RPL24 knockdown altered the expression of multiple miRs, and RPL24 immunoprecipitation indicated that up- or down-regulation of the mature miRs depended on whether their precursors bind RPL24 directly. Finally, further tests showed that RPL24 interacts directly with DDX5, a component of the large microprocessor complex, to inhibit miR processing. Our findings reveal that RPL24, which has previously been shown to play a role in miR processing in Arabidopsis thaliana, has a similar evolutionarily conserved function in miR biogenesis in mammals. We thus characterize a novel extra-ribosomal role of RPL24 in primate miR regulation.
Keywords: DDX5; Hsa-miR-608; Pre-miRs; Pri-miRs; RPL24; miR processing.
© 2024. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures

References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
