

Pro-inflammatory cytokines stimulate CFTR-dependent anion secretion in pancreatic ductal epithelium
- PMID: 38262945
- PMCID: PMC10807165
- DOI: 10.1186/s11658-024-00537-1
Pro-inflammatory cytokines stimulate CFTR-dependent anion secretion in pancreatic ductal epithelium
Abstract
Background: Loss of CFTR-dependent anion and fluid secretion in the ducts of the exocrine pancreas is thought to contribute to the development of pancreatitis, but little is known about the impact of inflammation on ductal CFTR function. Here we used adult stem cell-derived cell cultures (organoids) obtained from porcine pancreas to evaluate the effects of pro-inflammatory cytokines on CFTR function.
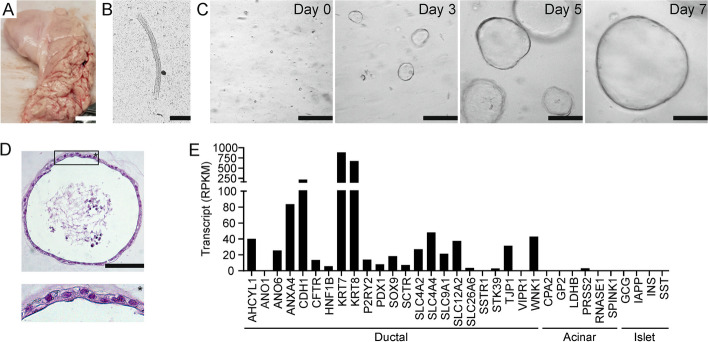
Methods: Organoids were cultured from porcine pancreas and used to prepare ductal epithelial monolayers. Monolayers were characterized by immunocytochemistry. Epithelial bicarbonate and chloride secretion, and the effect of IL-1β, IL-6, IFN-γ, and TNF-α on CFTR function was assessed by electrophysiology.
Results: Immunolocalization of ductal markers, including CFTR, keratin 7, and zonula occludens 1, demonstrated that organoid-derived cells formed a highly polarized epithelium. Stimulation by secretin or VIP triggered CFTR-dependent anion secretion across epithelial monolayers, whereas purinergic receptor stimulation by UTP, elicited CFTR-independent anion secretion. Most of the anion secretory response was attributable to bicarbonate transport. The combination of IL-1β, IL-6, IFN-γ, and TNF-α markedly enhanced CFTR expression and anion secretion across ductal epithelial monolayers, whereas these cytokines had little effect when tested separately. Although TNF-α triggered apoptotic signaling, epithelial barrier function was not significantly affected by cytokine exposure.
Conclusions: Pro-inflammatory cytokines enhance CFTR-dependent anion secretion across pancreatic ductal epithelium. We propose that up-regulation of CFTR in the early stages of the inflammatory response, may serve to promote the removal of pathogenic stimuli from the ductal tree, and limit tissue injury.
Keywords: CFTR; Cystic fibrosis; Cytokines; Epithelial ion transport; Organoid; Pancreatitis.
© 2024. The Author(s).
Conflict of interest statement
The authors declare that no competing interest exists.
Figures





Similar articles
-
Development of a polarized pancreatic ductular cell epithelium for physiological studies.J Appl Physiol (1985). 2018 Jul 1;125(1):97-106. doi: 10.1152/japplphysiol.00043.2018. Epub 2018 Mar 8. J Appl Physiol (1985). 2018. PMID: 29517421 Free PMC article.
-
Rescue of chloride and bicarbonate transport by elexacaftor-ivacaftor-tezacaftor in organoid-derived CF intestinal and cholangiocyte monolayers.J Cyst Fibros. 2022 May;21(3):537-543. doi: 10.1016/j.jcf.2021.12.006. Epub 2021 Dec 23. J Cyst Fibros. 2022. PMID: 34922851
-
Pancreatic duct organoid swelling is chloride-dependent.J Cyst Fibros. 2024 Jan;23(1):169-171. doi: 10.1016/j.jcf.2023.08.003. Epub 2023 Aug 24. J Cyst Fibros. 2024. PMID: 37633792 Free PMC article.
-
Bicarbonate Transport in Cystic Fibrosis and Pancreatitis.Cells. 2021 Dec 24;11(1):54. doi: 10.3390/cells11010054. Cells. 2021. PMID: 35011616 Free PMC article. Review.
-
Physiology and pathophysiology of bicarbonate secretion by pancreatic duct epithelium.Nagoya J Med Sci. 2012 Feb;74(1-2):1-18. Nagoya J Med Sci. 2012. PMID: 22515107 Free PMC article. Review.
Cited by
-
CFTR Modulators Counteract F508del CFTR Functional Defects in a Pancreatic Epithelial Model of Cystic Fibrosis.Life (Basel). 2025 Aug 19;15(8):1315. doi: 10.3390/life15081315. Life (Basel). 2025. PMID: 40868962 Free PMC article.
References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
