

Integrated compact regulators of protein activity enable control of signaling pathways and genome-editing in vivo
- PMID: 38263404
- PMCID: PMC10805712
- DOI: 10.1038/s41421-023-00632-1
Integrated compact regulators of protein activity enable control of signaling pathways and genome-editing in vivo
Abstract
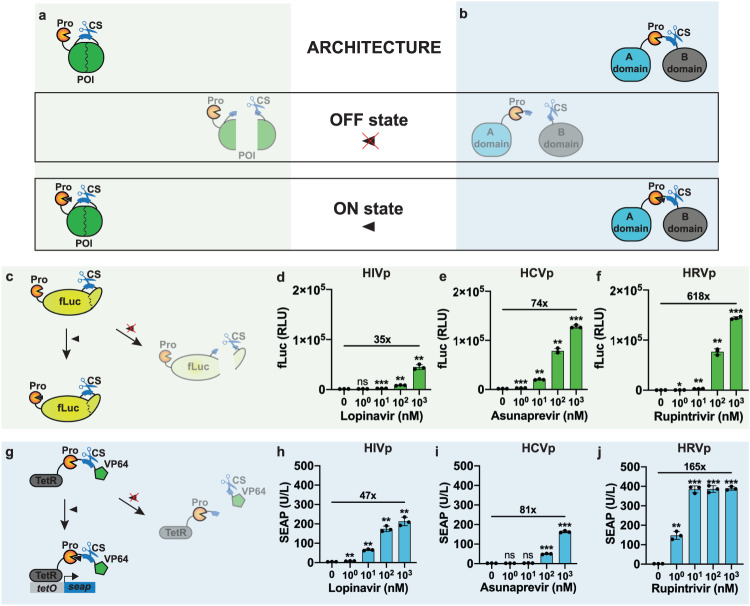
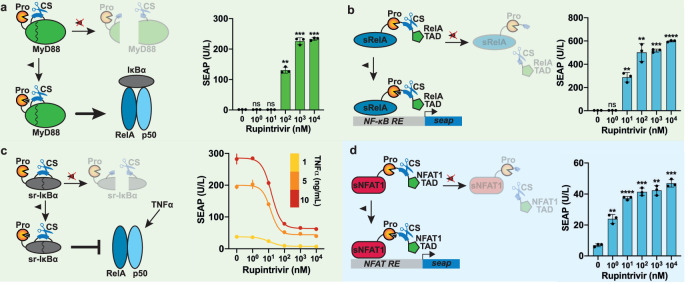
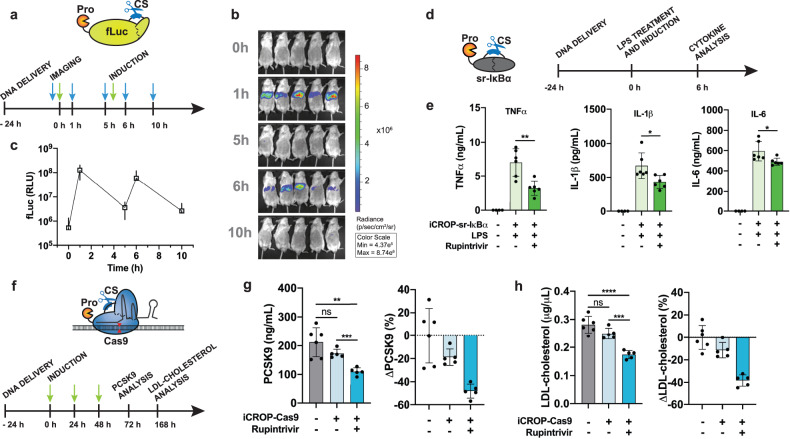
Viral proteases and clinically safe inhibitors were employed to build integrated compact regulators of protein activity (iCROP) for post-translational regulation of functional proteins by tunable proteolytic activity. In the absence of inhibitor, the co-localized/fused protease cleaves a target peptide sequence introduced in an exposed loop of the protein of interest, irreversibly fragmenting the protein structure and destroying its functionality. We selected three proteases and demonstrated the versatility of the iCROP framework by validating it to regulate the functional activity of ten different proteins. iCROP switches can be delivered either as mRNA or DNA, and provide rapid actuation kinetics with large induction ratios, while remaining strongly suppressed in the off state without inhibitor. iCROPs for effectors of the NF-κB and NFAT signaling pathways were assembled and confirmed to enable precise activation/inhibition of downstream events in response to protease inhibitors. In lipopolysaccharide-treated mice, iCROP-sr-IκBα suppressed cytokine release ("cytokine storm") by rescuing the activity of IκBα, which suppresses NF-κB signaling. We also constructed compact inducible CRISPR-(d)Cas9 variants and showed that iCROP-Cas9-mediated knockout of the PCSK9 gene in the liver lowered blood LDL-cholesterol levels in mice. iCROP-based protein switches will facilitate protein-level regulation in basic research and translational applications.
© 2023. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures


References
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
