

Impaired STING Activation Due to a Variant in the E3 Ubiquitin Ligase AMFR in a Patient with Severe VZV Infection and Hemophagocytic Lymphohistiocytosis
- PMID: 38277122
- PMCID: PMC10817851
- DOI: 10.1007/s10875-024-01653-5
Impaired STING Activation Due to a Variant in the E3 Ubiquitin Ligase AMFR in a Patient with Severe VZV Infection and Hemophagocytic Lymphohistiocytosis
Abstract
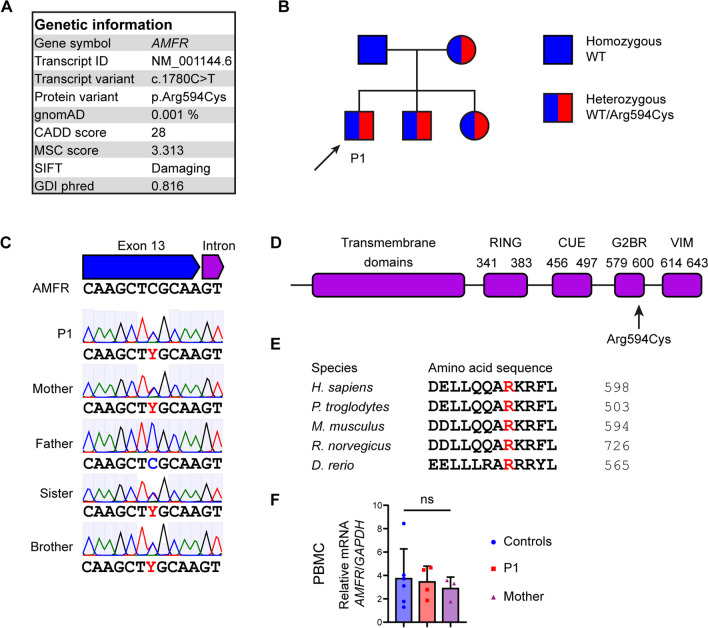
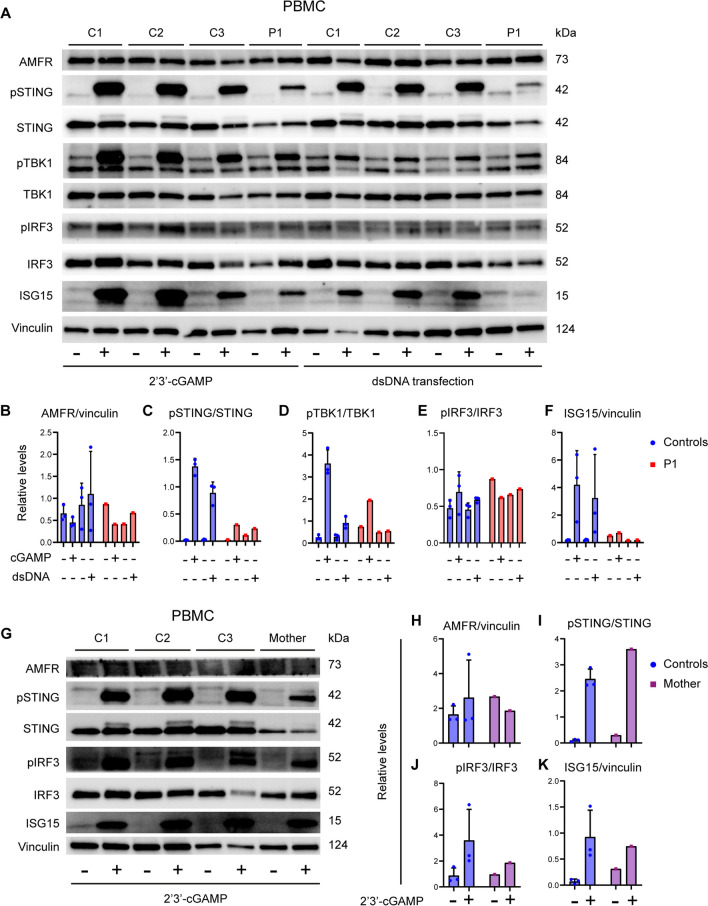
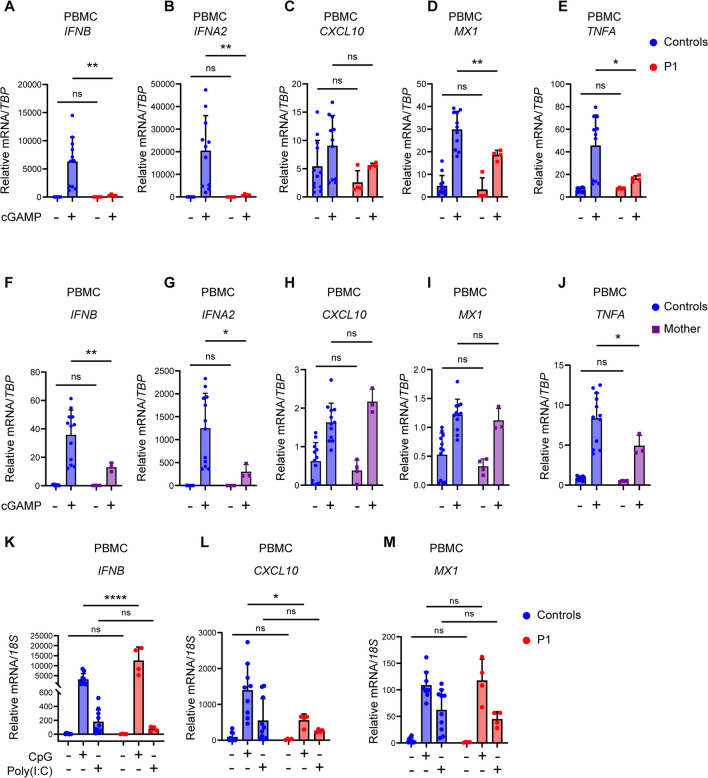
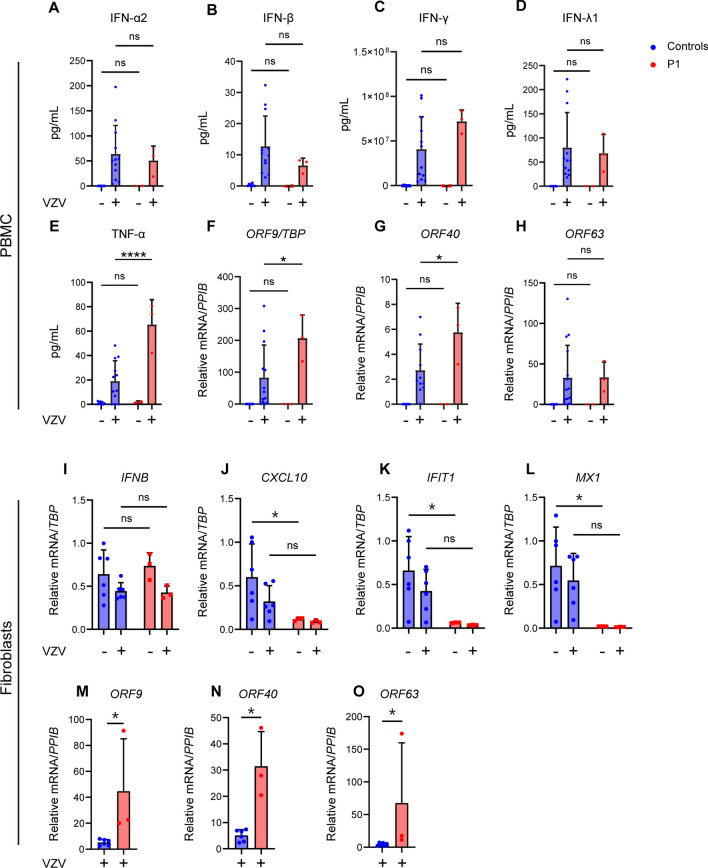
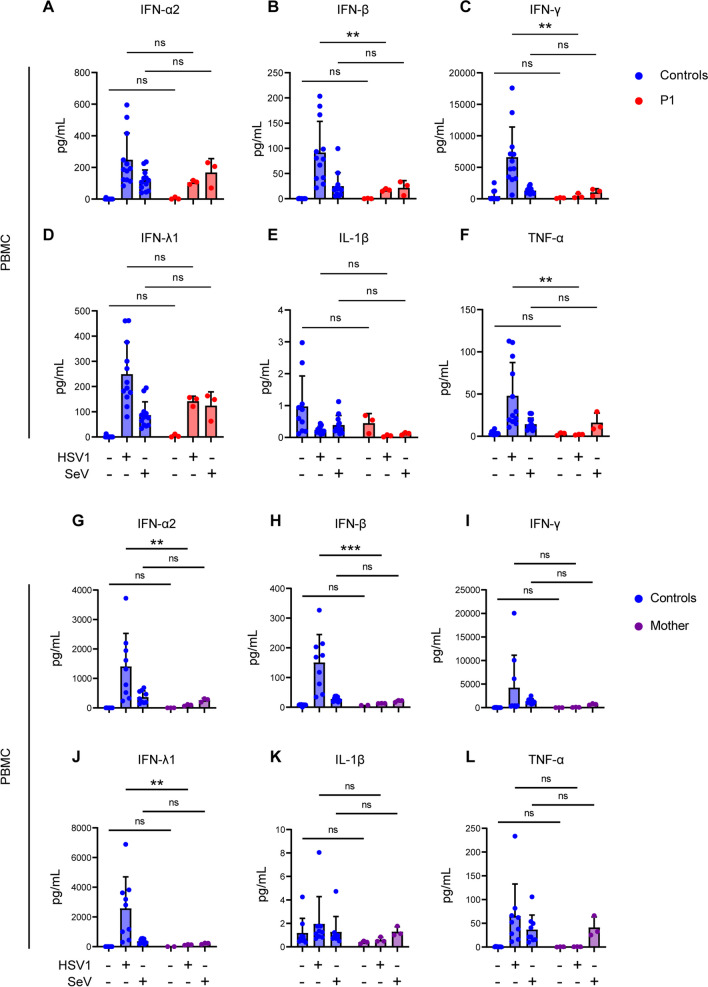
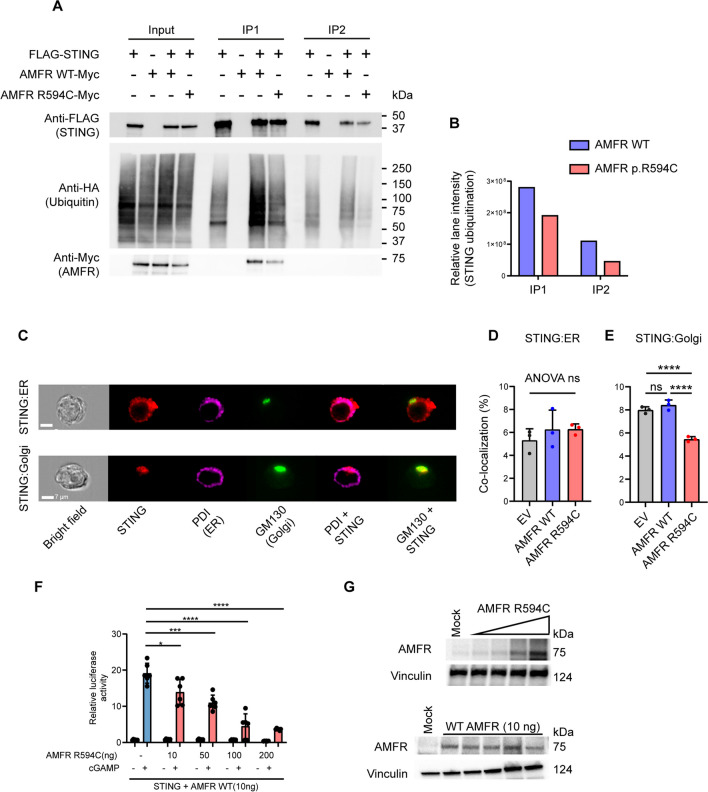
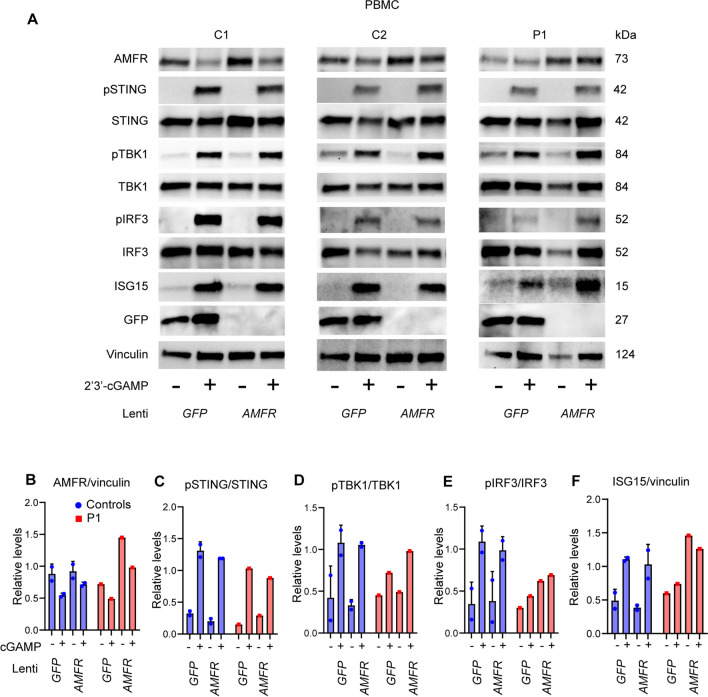
Varicella zoster virus (VZV) is a neurotropic alphaherpesvirus exclusively infecting humans, causing two distinct pathologies: varicella (chickenpox) upon primary infection and herpes zoster (shingles) following reactivation. In susceptible individuals, VZV can give rise to more severe clinical manifestations, including disseminated infection, pneumonitis, encephalitis, and vasculopathy with stroke. Here, we describe a 3-year-old boy in whom varicella followed a complicated course with thrombocytopenia, hemorrhagic and necrotic lesions, pneumonitis, and intermittent encephalopathy. Hemophagocytic lymphohistiocytosis (HLH) was strongly suspected and as the condition deteriorated, HLH therapy was initiated. Although the clinical condition improved, longstanding hemophagocytosis followed despite therapy. We found that the patient carries a rare monoallelic variant in autocrine motility factor receptor (AMFR), encoding a ubiquitin ligase involved in innate cytosolic DNA sensing and interferon (IFN) production through the cyclic GMP-AMP synthase-stimulator of IFN genes (cGAS-STING) pathway. Peripheral blood mononuclear cells (PBMCs) from the patient exhibited impaired signaling downstream of STING in response dsDNA and 2'3'-cGAMP, agonists of cGAS and STING, respectively, and fibroblasts from the patient showed impaired type I IFN responses and significantly increased VZV replication. Overexpression of the variant AMFR R594C resulted in decreased K27-linked STING ubiquitination compared to WT AMFR. Moreover, ImageStream technology revealed reduced STING trafficking from ER to Golgi in cells expressing the patient AMFR R594C variant. This was supported by a dose-dependent dominant negative effect of expression of the patient AMFR variant as measured by IFN-β reporter gene assay. Finally, lentiviral transduction with WT AMFR partially reconstituted 2'3'-cGAMP-induced STING-mediated signaling and ISG expression in patient PBMCs. This work links defective AMFR-STING signaling to severe VZV disease and hyperinflammation and suggests a direct role for cGAS-STING in the control of viral infections in humans. In conclusion, we describe a novel genetic etiology of severe VZV disease in childhood, also representing the first inborn error of immunity related to a defect in the cGAS-STING pathway.
Keywords: AMFR; HLH; ISG; Interferon; STING; Ubiquitin ligase; VZV.
© 2024. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Research Materials
