

Sulfated bile acid is a host-derived ligand for MAIT cells
- PMID: 38277465
- PMCID: PMC11147531
- DOI: 10.1126/sciimmunol.ade6924
Sulfated bile acid is a host-derived ligand for MAIT cells
Abstract
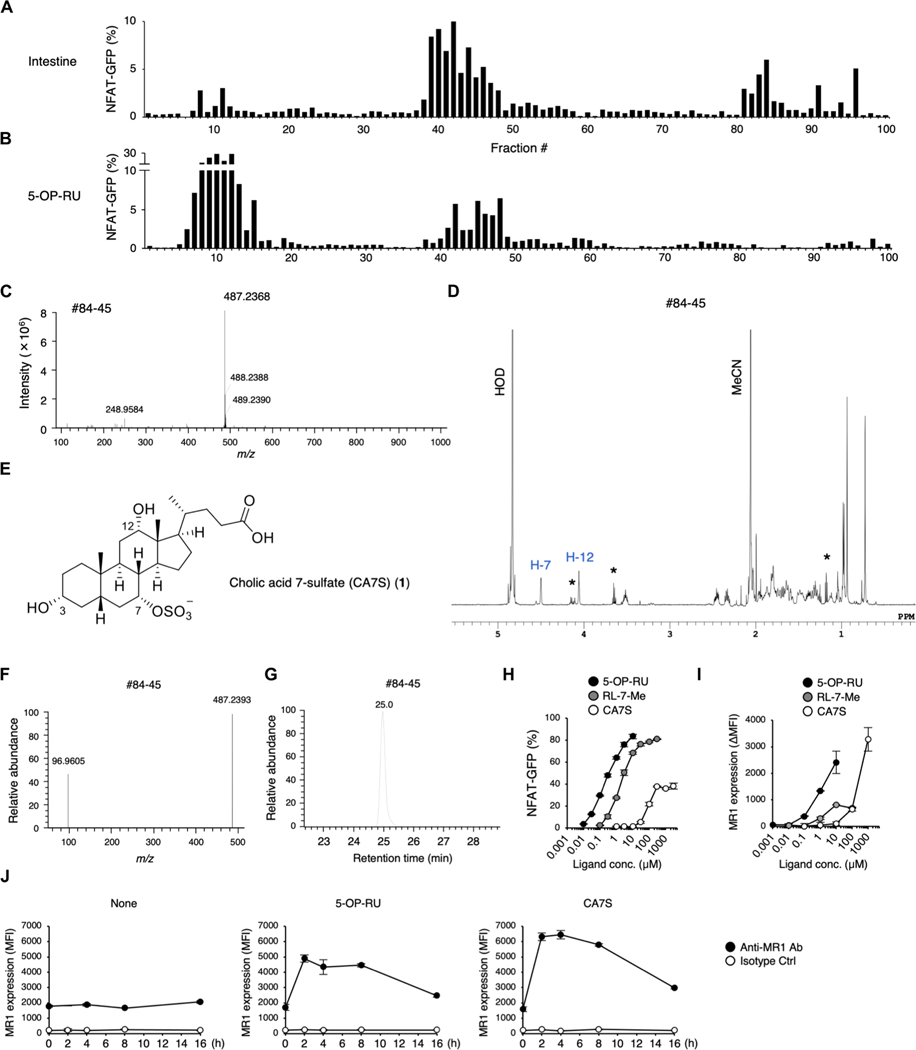
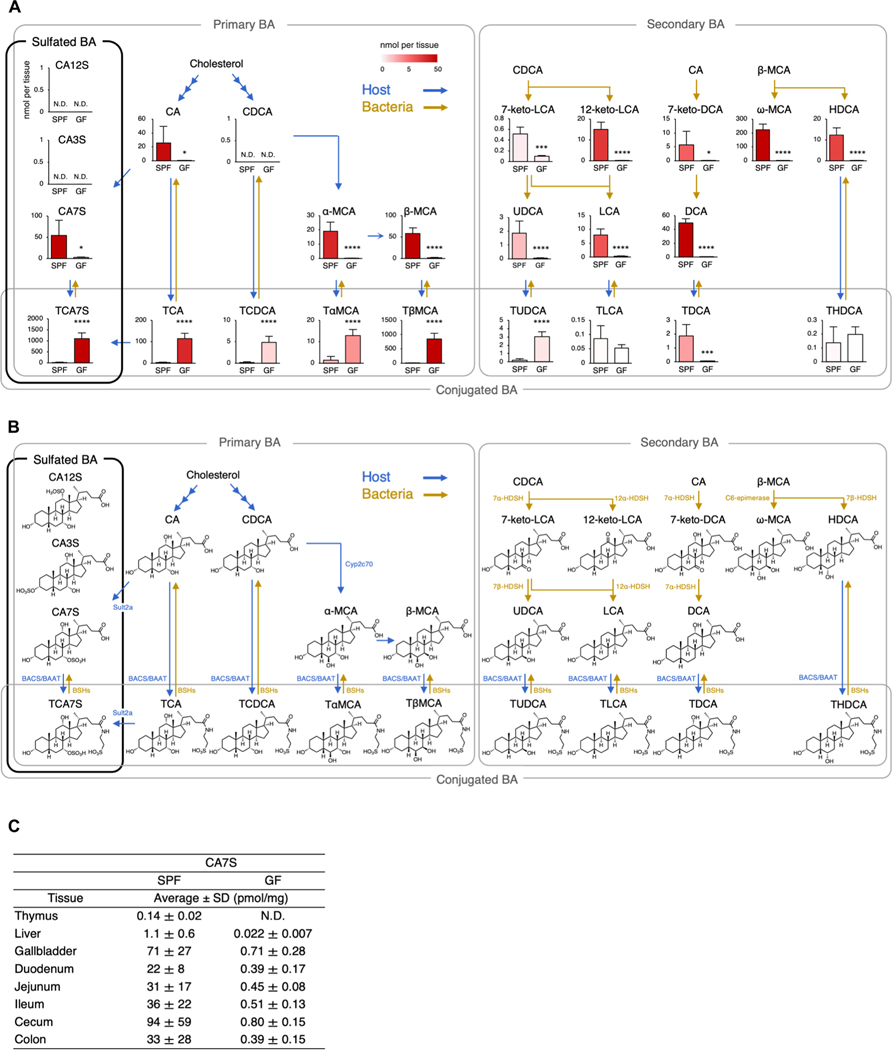
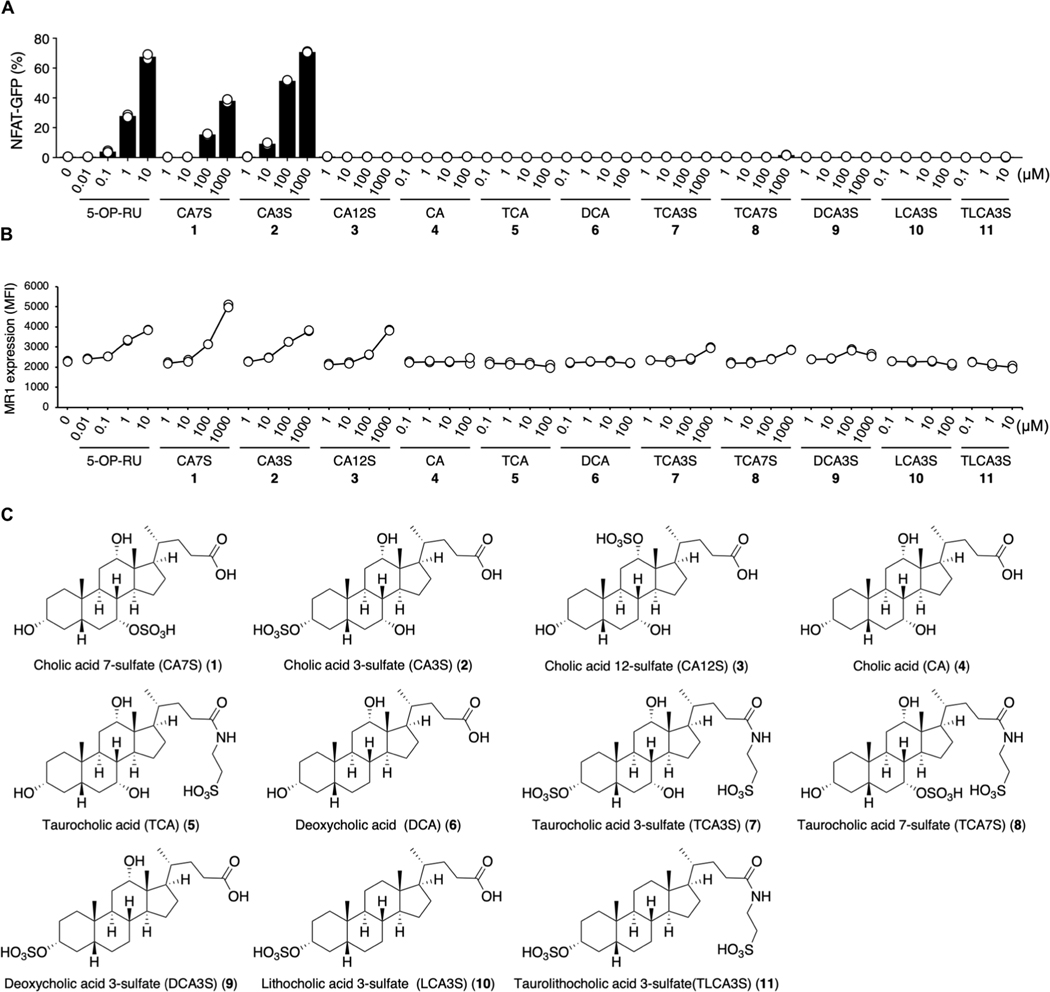
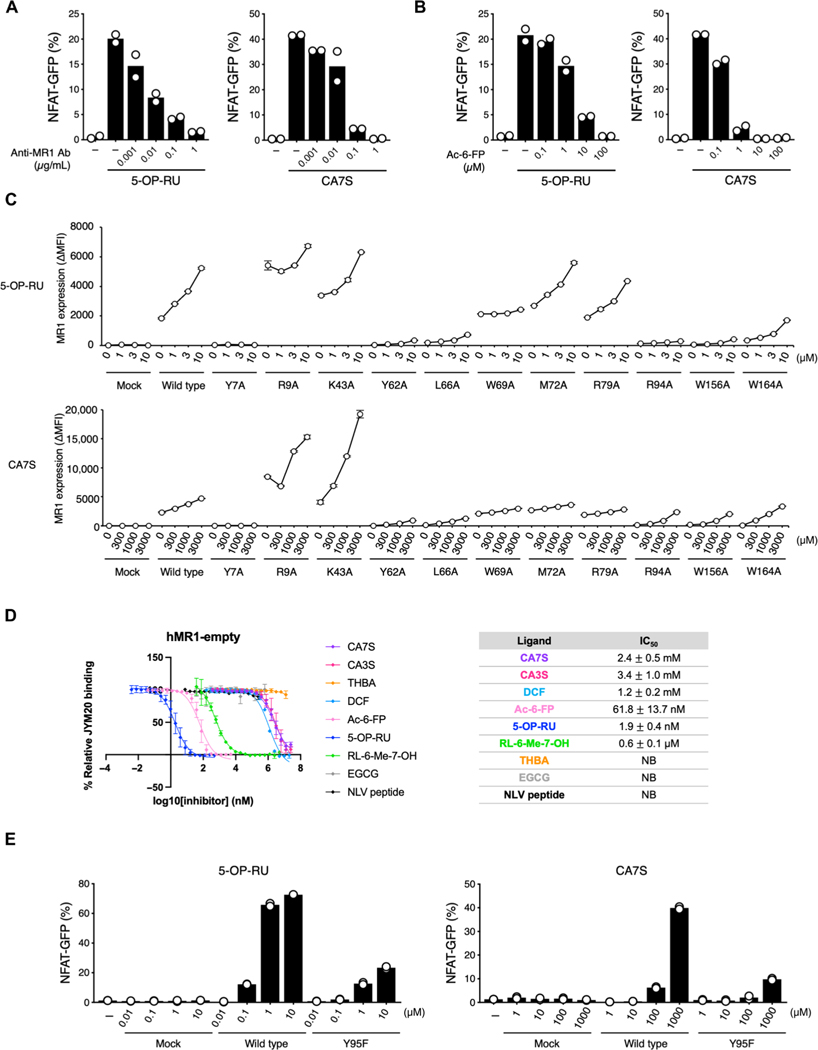
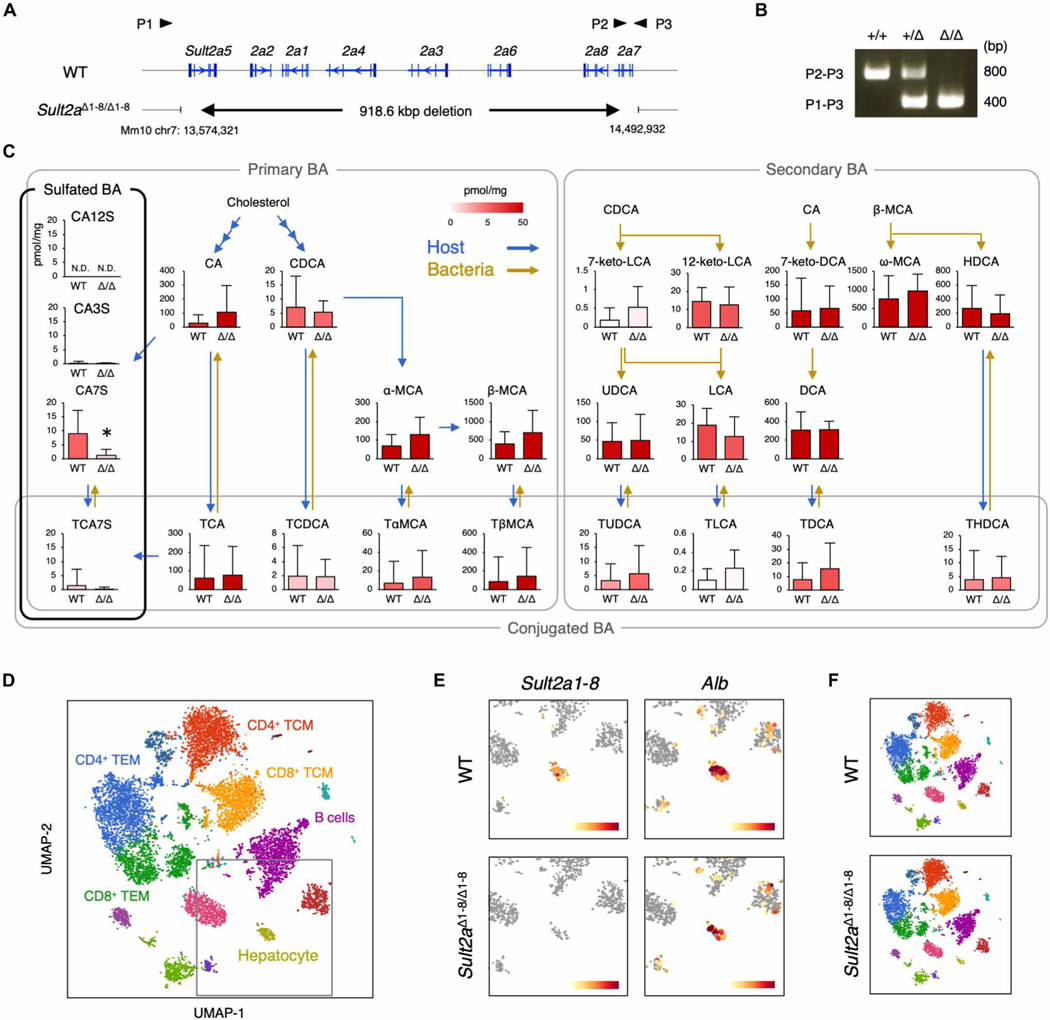
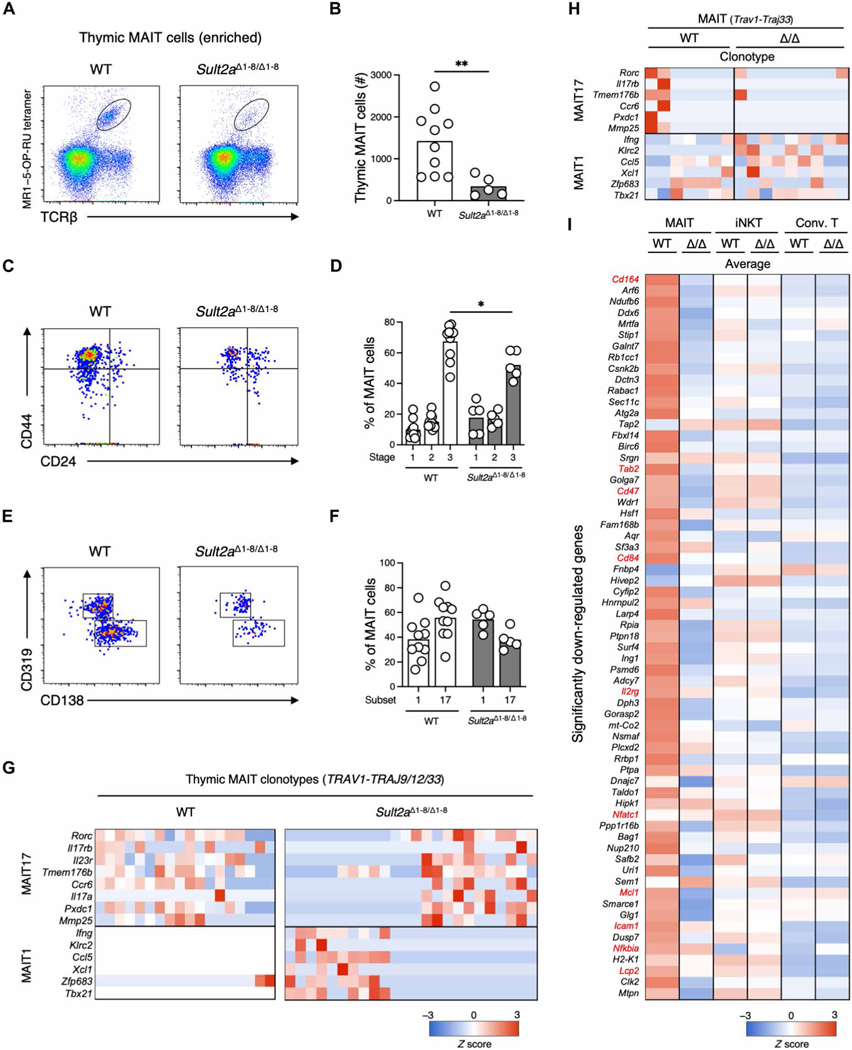
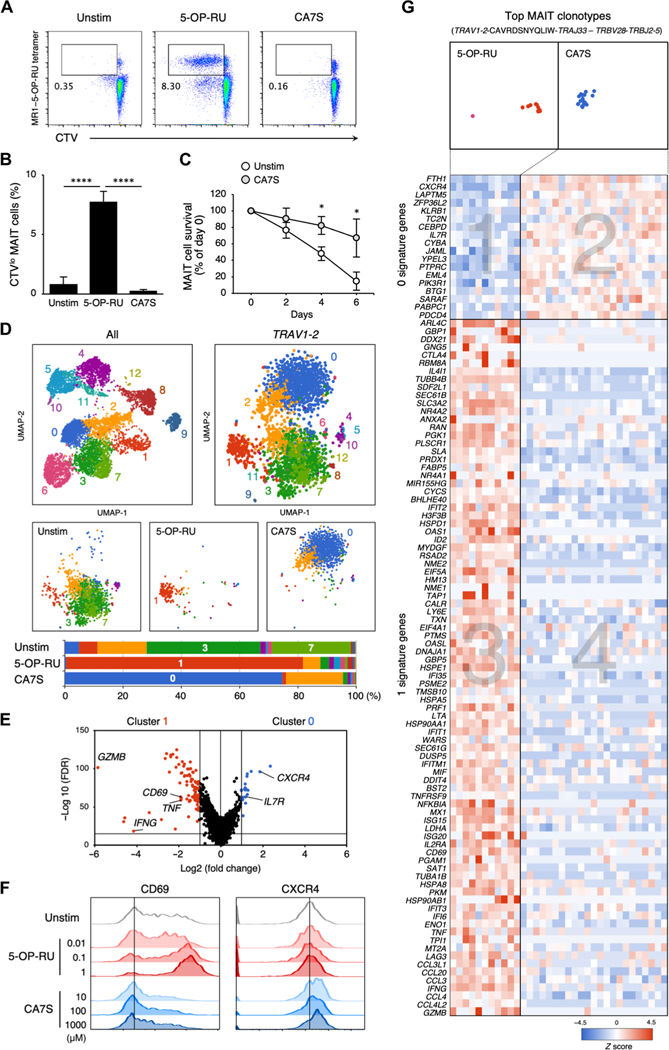
Mucosal-associated invariant T (MAIT) cells are innate-like T cells that recognize bacterial riboflavin-based metabolites as activating antigens. Although MAIT cells are found in tissues, it is unknown whether any host tissue-derived antigens exist. Here, we report that a sulfated bile acid, cholic acid 7-sulfate (CA7S), binds the nonclassical MHC class I protein MR1 and is recognized by MAIT cells. CA7S is a host-derived metabolite whose levels were reduced by more than 98% in germ-free mice. Deletion of the sulfotransferase 2a family of enzymes (Sult2a1-8) responsible for CA7S synthesis reduced the number of thymic MAIT cells in mice. Moreover, recognition of CA7S induced MAIT cell survival and the expression of a homeostatic gene signature. By contrast, recognition of a previously described foreign antigen, 5-(2-oxopropylideneamino)-6-d-ribitylaminouracil (5-OP-RU), drove MAIT cell proliferation and the expression of inflammatory genes. Thus, CA7S is an endogenous antigen for MAIT cells, which promotes their development and function.
Conflict of interest statement
Figures
References
-
- Godfrey DI, Koay HF, McCluskey J, Gherardin NA, The biology and functional importance of MAIT cells. Nat. Immunol 20, 1110–1128 (2019). - PubMed
-
- Legoux F, Salou M, Lantz O, MAIT cell development and functions: The microbial connection. Immunity 53, 710–723 (2020). - PubMed
-
- Treiner E, Duban L, Bahram S, Radosavljevic M, Wanner V, Tilloy F, Affaticati P, Gilfillan S, Lantz O, Addendum: Selection of evolutionarily conserved mucosal-associated invariant T cells by MR1. Nature 423, 1018 (2003). - PubMed
-
- Kjer-Nielsen L, Patel O, Corbett AJ, Le Nours J, Meehan B, Liu L, Bhati M, Chen Z, Kostenko L, Reantragoon R, Williamson NA, Purcell AW, Dudek NL, McConville MJ, O’Hair RAJ, Khairallah GN, Godfrey DI, Fairlie DP, Rossjohn J, McCluskey J, MR1 presents microbial vitamin B metabolites to MAIT cells. Nature 491, 717–723 (2012). - PubMed
-
- Corbett AJ, Eckle SBG, Birkinshaw RW, Liu L, Patel O, Mahony J, Chen Z, Reantragoon R, Meehan B, Cao H, Williamson NA, Strugnell RA, Van Sinderen D, Mak JYW, Fairlie DP, Kjer-Nielsen L, Rossjohn J, McCluskey J, T-cell activation by transitory neo-antigens derived from distinct microbial pathways. Nature 509, 361–365 (2014). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
