

Mis-spliced transcripts generate de novo proteins in TDP-43-related ALS/FTD
- PMID: 38277467
- PMCID: PMC11325748
- DOI: 10.1126/scitranslmed.adg7162
Mis-spliced transcripts generate de novo proteins in TDP-43-related ALS/FTD
Abstract
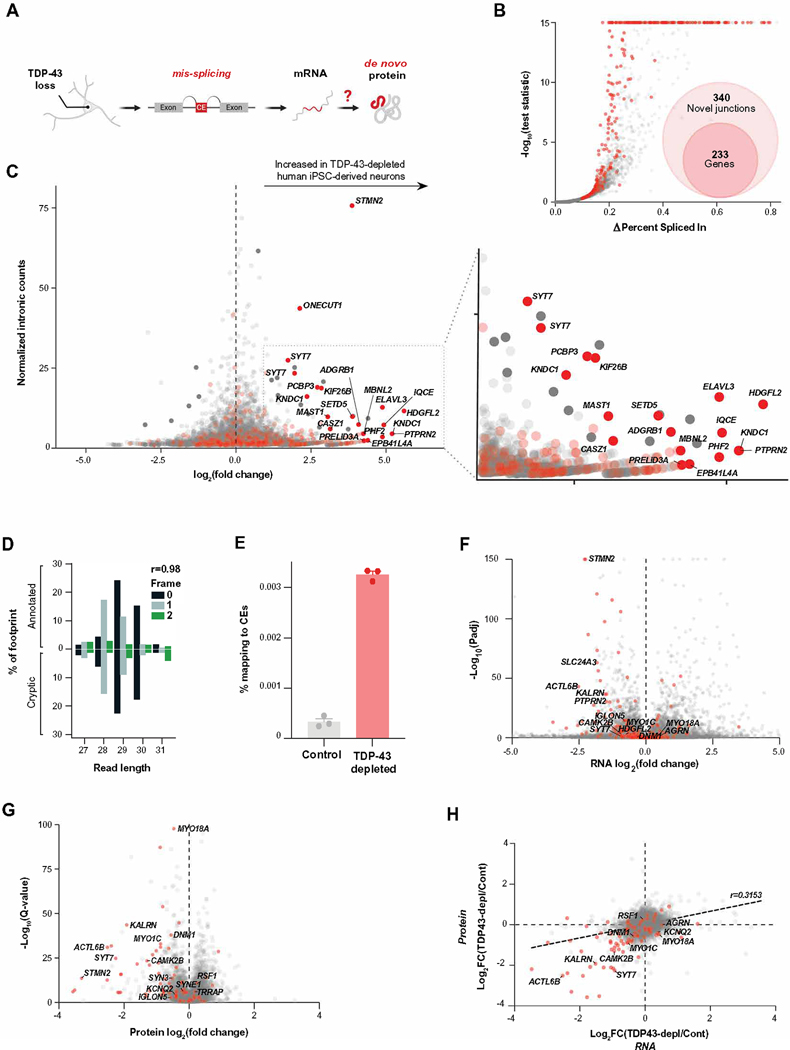
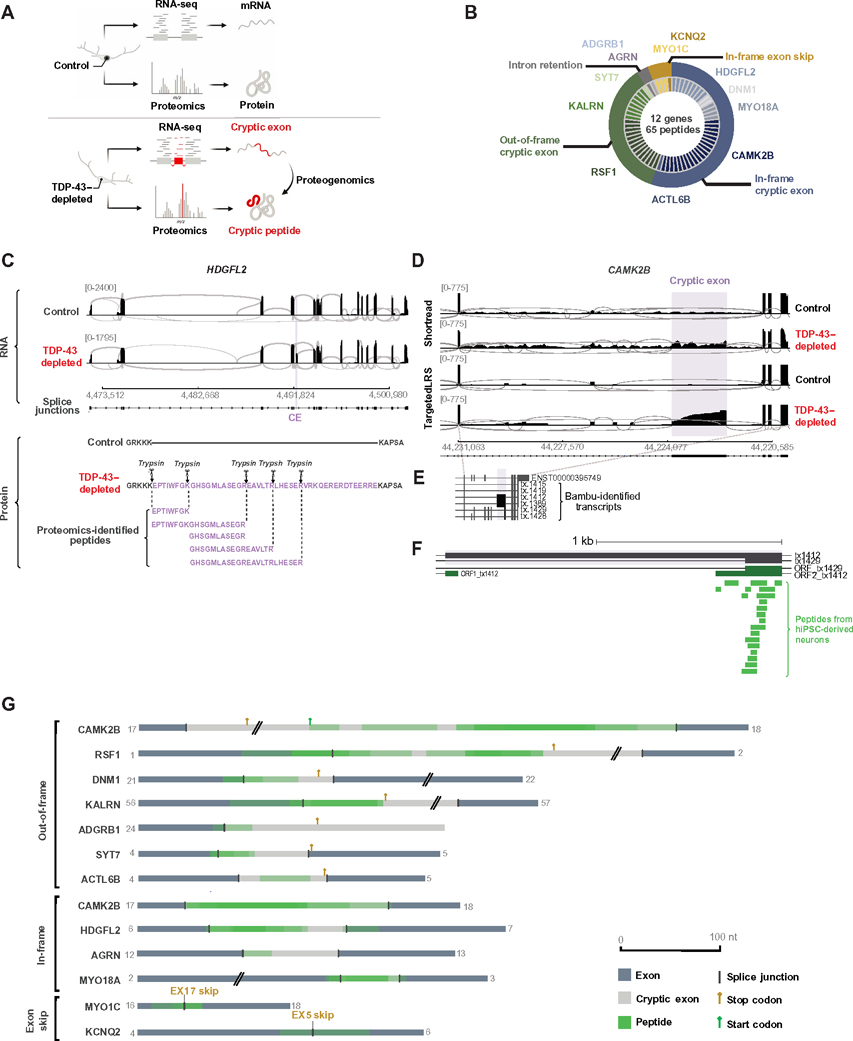
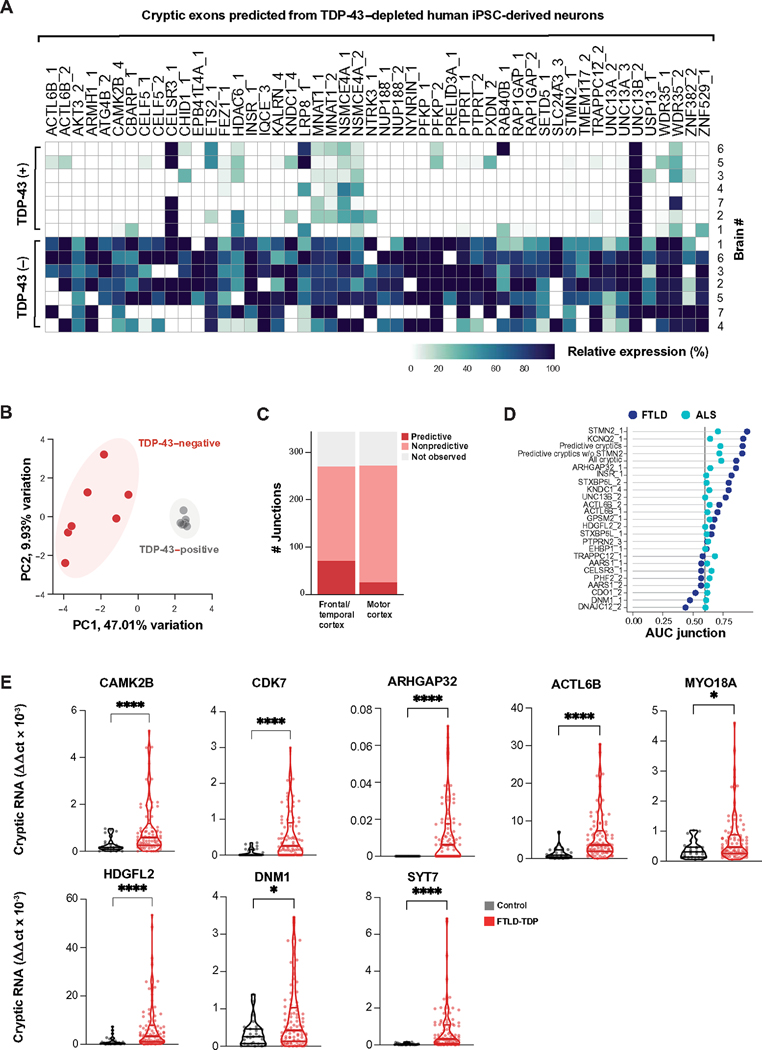
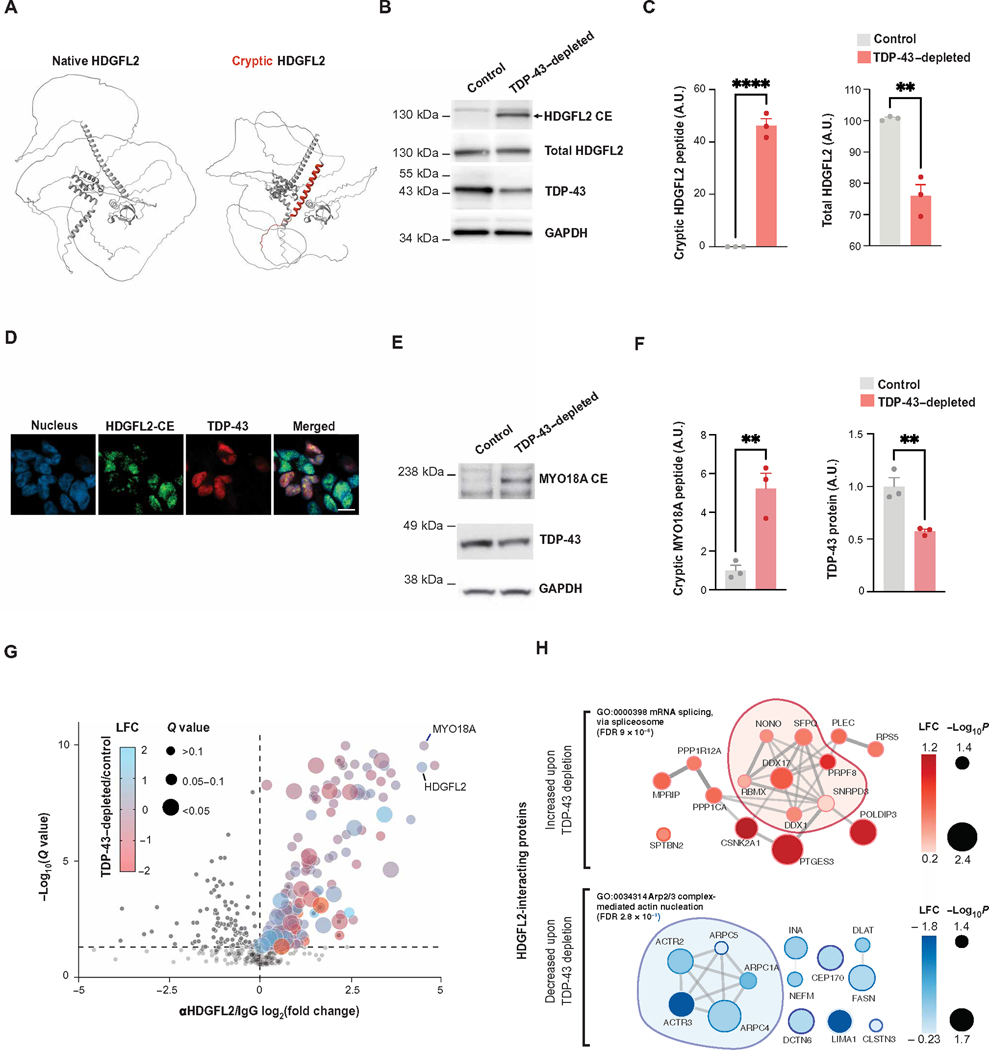
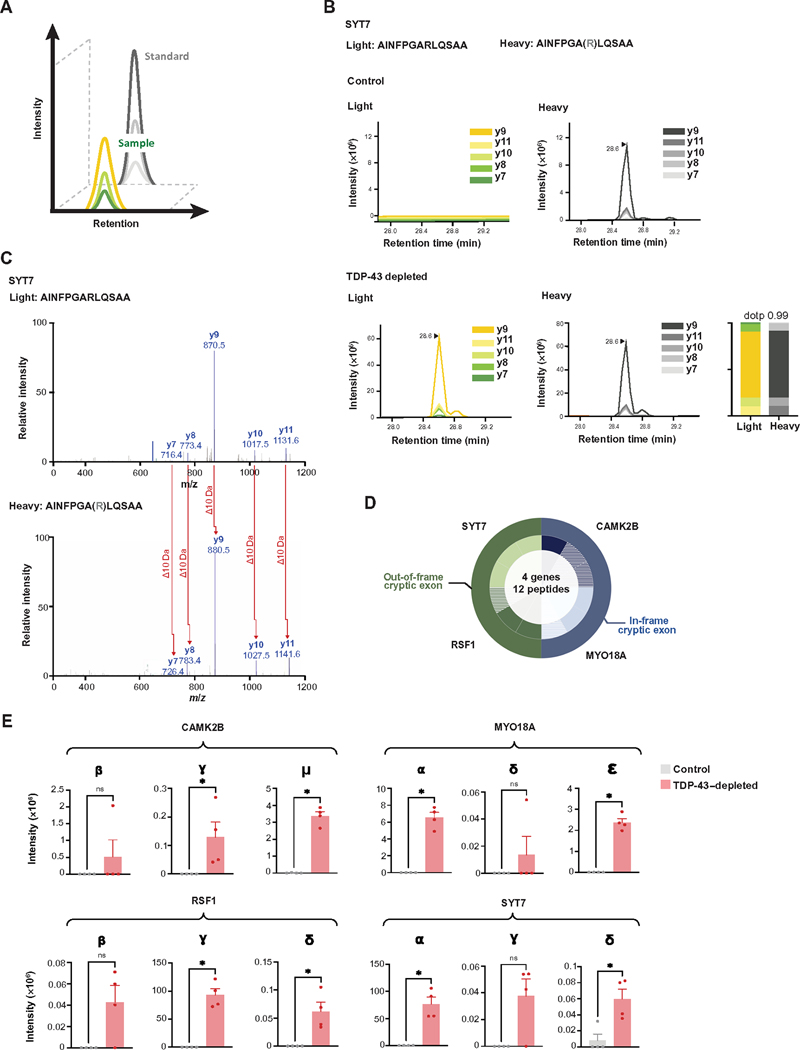
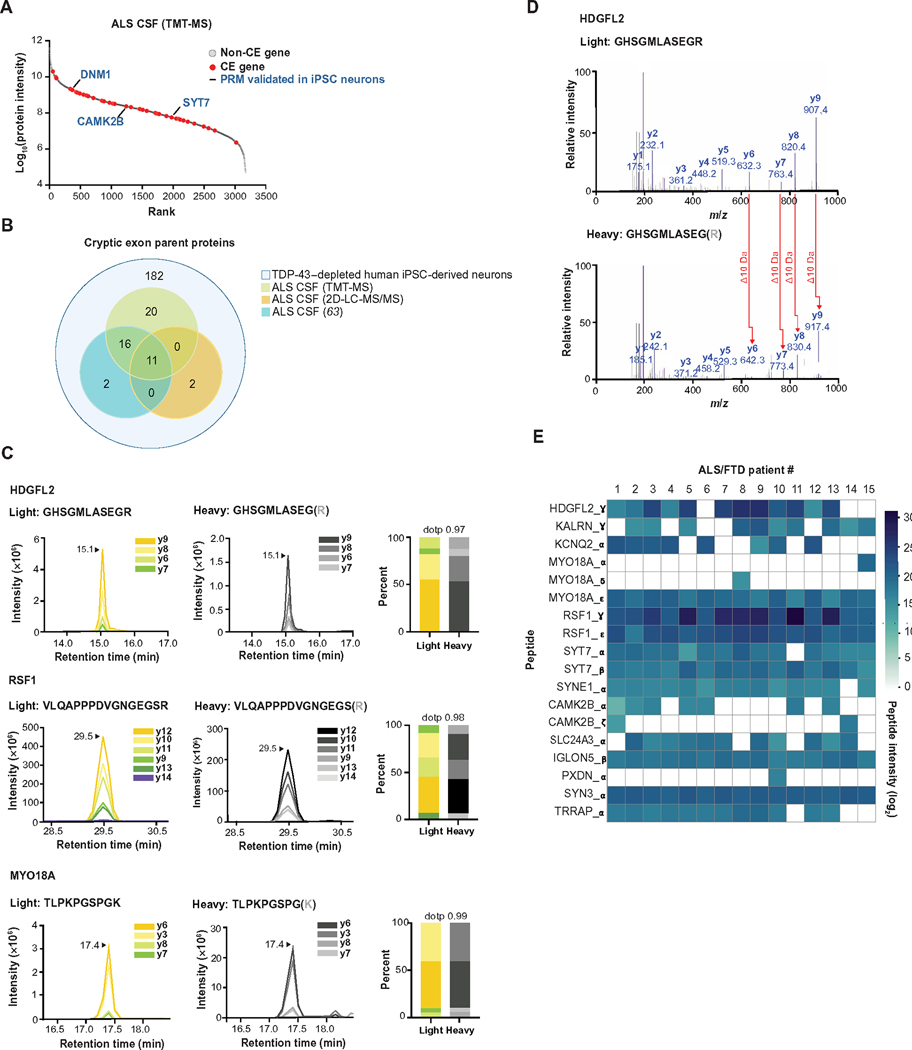
Functional loss of TDP-43, an RNA binding protein genetically and pathologically linked to amyotrophic lateral sclerosis (ALS) and frontotemporal dementia (FTD), leads to the inclusion of cryptic exons in hundreds of transcripts during disease. Cryptic exons can promote the degradation of affected transcripts, deleteriously altering cellular function through loss-of-function mechanisms. Here, we show that mRNA transcripts harboring cryptic exons generated de novo proteins in TDP-43-depleted human iPSC-derived neurons in vitro, and de novo peptides were found in cerebrospinal fluid (CSF) samples from patients with ALS or FTD. Using coordinated transcriptomic and proteomic studies of TDP-43-depleted human iPSC-derived neurons, we identified 65 peptides that mapped to 12 cryptic exons. Cryptic exons identified in TDP-43-depleted human iPSC-derived neurons were predictive of cryptic exons expressed in postmortem brain tissue from patients with TDP-43 proteinopathy. These cryptic exons produced transcript variants that generated de novo proteins. We found that the inclusion of cryptic peptide sequences in proteins altered their interactions with other proteins, thereby likely altering their function. Last, we showed that 18 de novo peptides across 13 genes were present in CSF samples from patients with ALS/FTD spectrum disorders. The demonstration of cryptic exon translation suggests new mechanisms for ALS/FTD pathophysiology downstream of TDP-43 dysfunction and may provide a potential strategy to assay TDP-43 function in patient CSF.
Conflict of interest statement
Figures
Update of
-
Mis-spliced transcripts generate de novo proteins in TDP-43-related ALS/FTD.bioRxiv [Preprint]. 2023 Jan 23:2023.01.23.525149. doi: 10.1101/2023.01.23.525149. bioRxiv. 2023. Update in: Sci Transl Med. 2024 Feb 14;16(734):eadg7162. doi: 10.1126/scitranslmed.adg7162. PMID: 36747793 Free PMC article. Updated. Preprint.
References
-
- Neumann M, Tolnay M, Mackenzie IRA, The molecular basis of frontotemporal dementia. Expert Rev. Mol. Med. 11, e23 (2009). - PubMed
-
- Van Deerlin VM, Leverenz JB, Bekris LM, Bird TD, Yuan W, Elman LB, Clay D, Wood EMC, Chen-Plotkin AS, Martinez-Lage M, Steinbart E, McCluskey L, Grossman M, Neumann M, Wu IL, Yang WS, Kalb R, Galasko DR, Montine TJ, Trojanowski JQ, Lee VMY, Schellenberg GD, Yu CE, TARDBP mutations in amyotrophic lateral sclerosis with TDP-43 neuropathology: A genetic and histopathological analysis. Lancet Neurol. 7, 409–416 (2008). - PMC - PubMed
-
- Borroni B, Bonvicini C, Alberici A, Buratti E, Agosti C, Archetti S, Papetti A, Stuani C, Di Luca M, Gennarelli M, Padovani A, Mutation withinTARDBPleads to frontotemporal dementia without motor neuron disease. Hum. Mutat. 30, E974–E983 (2009). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- R35 NS097273/NS/NINDS NIH HHS/United States
- RF1 AG062171/AG/NIA NIH HHS/United States
- T32 GM136577/GM/NIGMS NIH HHS/United States
- MC_PC_MR/S022708/1/MRC_/Medical Research Council/United Kingdom
- U19 AG063911/AG/NIA NIH HHS/United States
- CC0102/WT_/Wellcome Trust/United Kingdom
- MR/J009482/1/MRC_/Medical Research Council/United Kingdom
- P30 AG062677/AG/NIA NIH HHS/United States
- U54 NS123743/NS/NINDS NIH HHS/United States
- ZIA NS003155/ImNIH/Intramural NIH HHS/United States
- MR/T046015/1/MRC_/Medical Research Council/United Kingdom
- MR/M008525/1/MRC_/Medical Research Council/United Kingdom
- MR/W005190/1/MRC_/Medical Research Council/United Kingdom
- FI2 GM142475/GM/NIGMS NIH HHS/United States
- MR/S006508/1/MRC_/Medical Research Council/United Kingdom
- MALASPINA/APR13/817-791/MNDA_/Motor Neurone Disease Association/United Kingdom
- HALLEGGER/OCT15/959-799/MNDA_/Motor Neurone Disease Association/United Kingdom
- RF1 AG062077/AG/NIA NIH HHS/United States
- MR/M023664/1/MRC_/Medical Research Council/United Kingdom
- P01 NS084974/NS/NINDS NIH HHS/United States
LinkOut - more resources
Full Text Sources
Medical
Miscellaneous
