

A fluid biomarker reveals loss of TDP-43 splicing repression in presymptomatic ALS-FTD
- PMID: 38278991
- PMCID: PMC10878965
- DOI: 10.1038/s41591-023-02788-5
A fluid biomarker reveals loss of TDP-43 splicing repression in presymptomatic ALS-FTD
Erratum in
-
Author Correction: A fluid biomarker reveals loss of TDP-43 splicing repression in presymptomatic ALS-FTD.Nat Med. 2024 May;30(5):1504. doi: 10.1038/s41591-024-02966-z. Nat Med. 2024. PMID: 38580817 Free PMC article. No abstract available.
Abstract
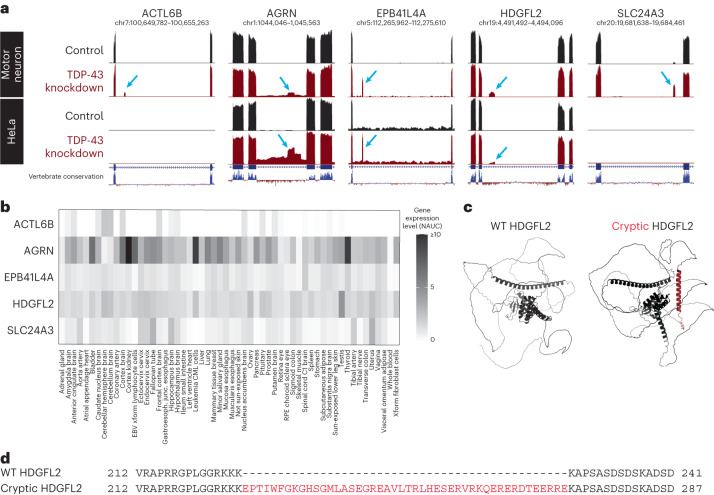
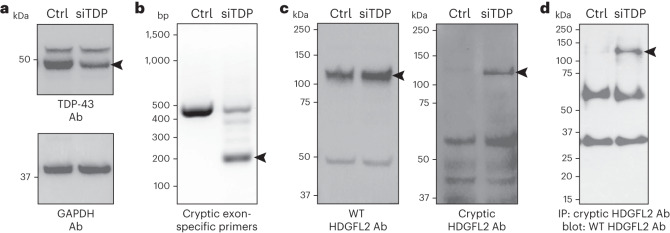
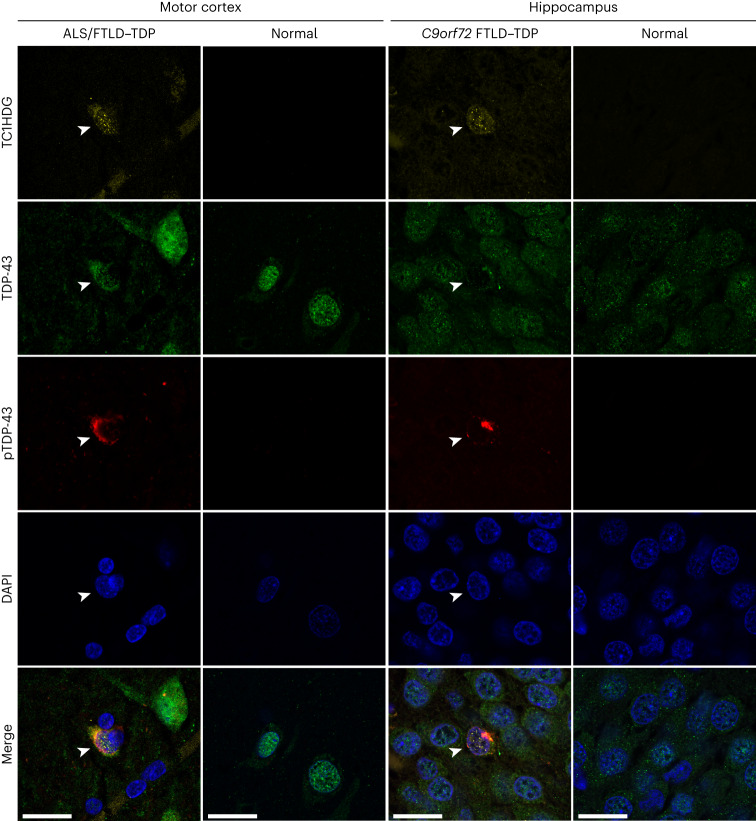
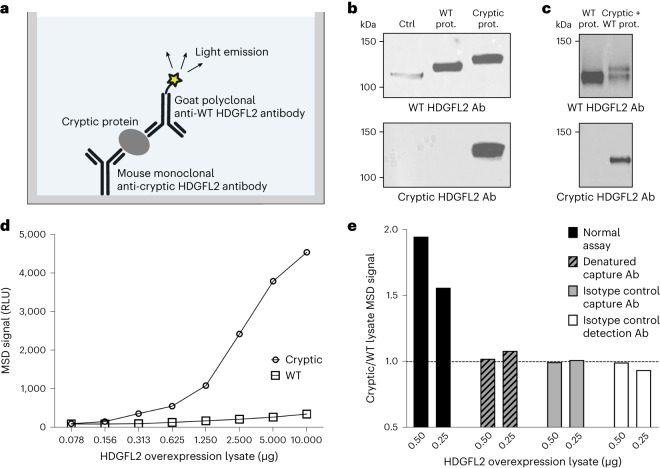
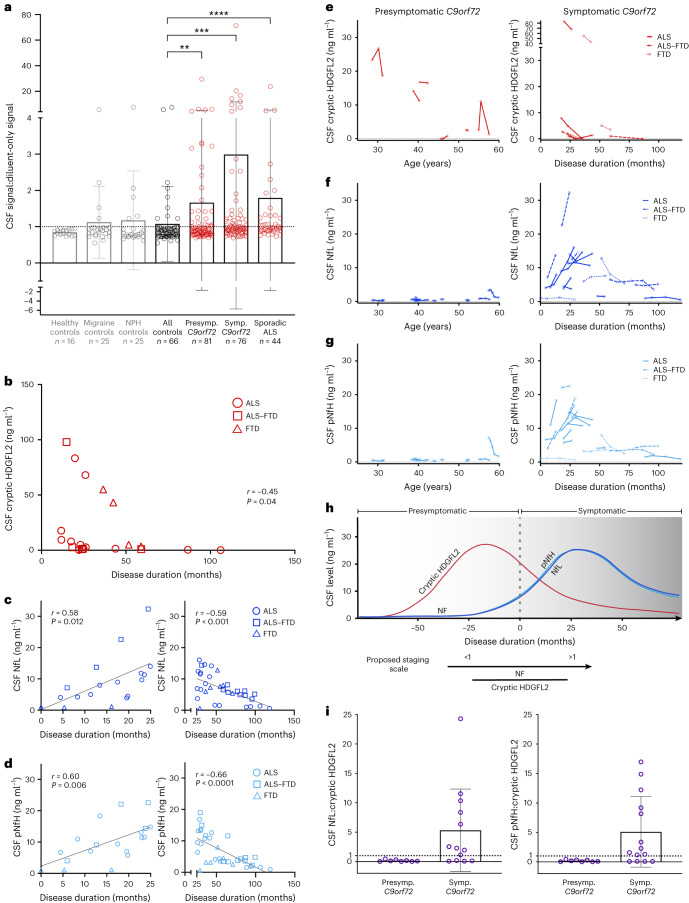
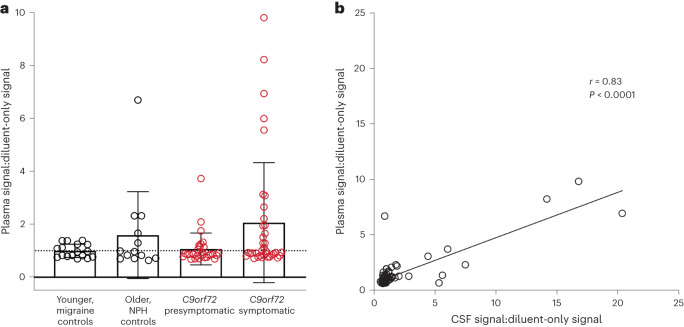
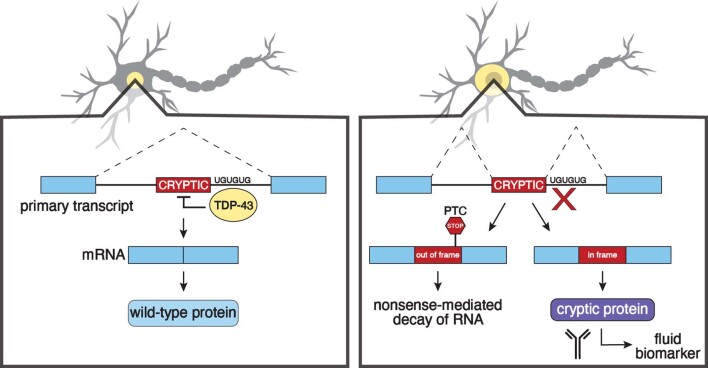
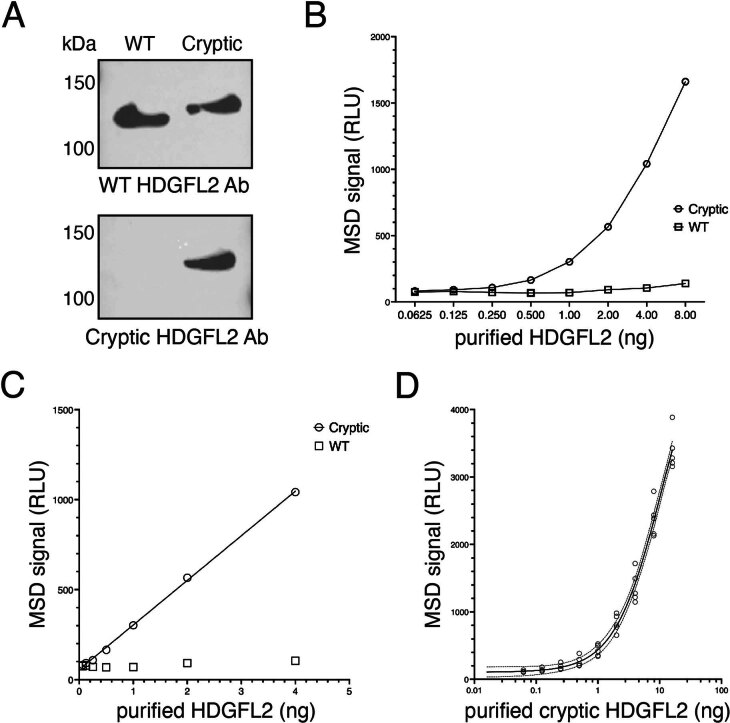
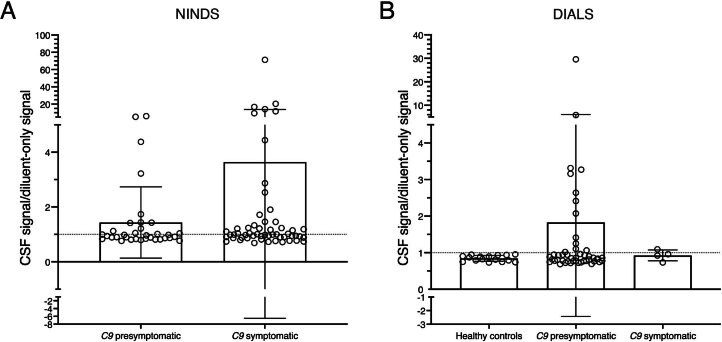
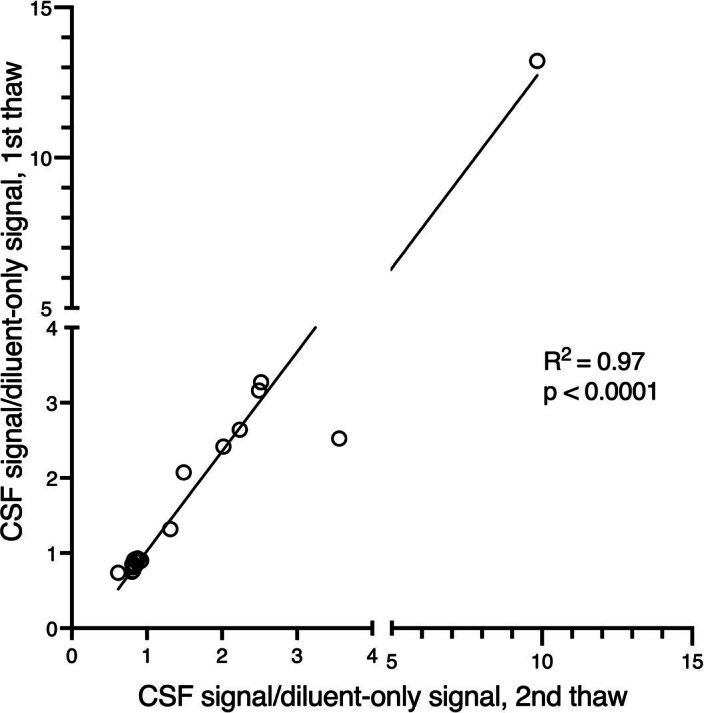
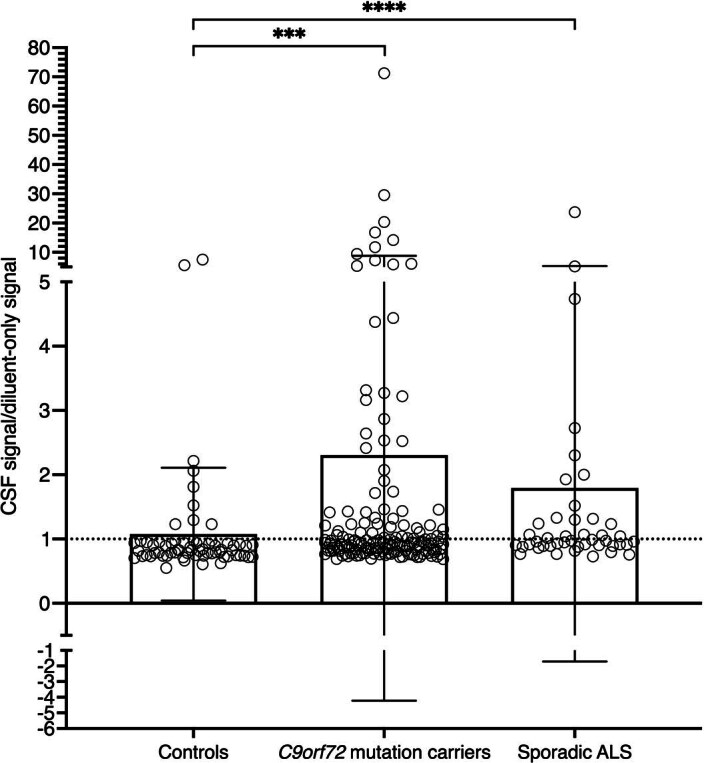
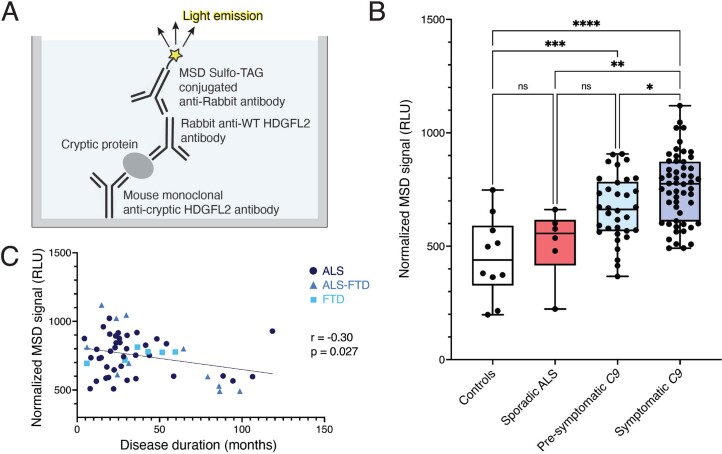
Although loss of TAR DNA-binding protein 43 kDa (TDP-43) splicing repression is well documented in postmortem tissues of amyotrophic lateral sclerosis (ALS) and frontotemporal dementia (FTD), whether this abnormality occurs during early-stage disease remains unresolved. Cryptic exon inclusion reflects loss of function of TDP-43, and thus detection of proteins containing cryptic exon-encoded neoepitopes in cerebrospinal fluid (CSF) or blood could reveal the earliest stages of TDP-43 dysregulation in patients. Here we use a newly characterized monoclonal antibody specific to a TDP-43-dependent cryptic epitope (encoded by the cryptic exon found in HDGFL2) to show that loss of TDP-43 splicing repression occurs in ALS-FTD, including in presymptomatic C9orf72 mutation carriers. Cryptic hepatoma-derived growth factor-like protein 2 (HDGFL2) accumulates in CSF at significantly higher levels in familial ALS-FTD and sporadic ALS compared with controls and is elevated earlier than neurofilament light and phosphorylated neurofilament heavy chain protein levels in familial disease. Cryptic HDGFL2 can also be detected in blood of individuals with ALS-FTD, including in presymptomatic C9orf72 mutation carriers, and accumulates at levels highly correlated with those in CSF. Our findings indicate that loss of TDP-43 cryptic splicing repression occurs early in disease progression, even presymptomatically, and that detection of the HDGFL2 cryptic neoepitope serves as a potential diagnostic biomarker for ALS, which should facilitate patient recruitment and measurement of target engagement in clinical trials.
© 2024. The Author(s).
Conflict of interest statement
The authors declare the following competing interests. J.P.L. and P.C.W. are inventors on a provisional patent application submitted by Johns Hopkins University that covers the usage of TDP-43-associated cryptic exon-derived neoepitopes as biomarkers. B.J.T. holds patents on the clinical testing and therapeutic intervention for the hexanucleotide repeat expansion of
Figures
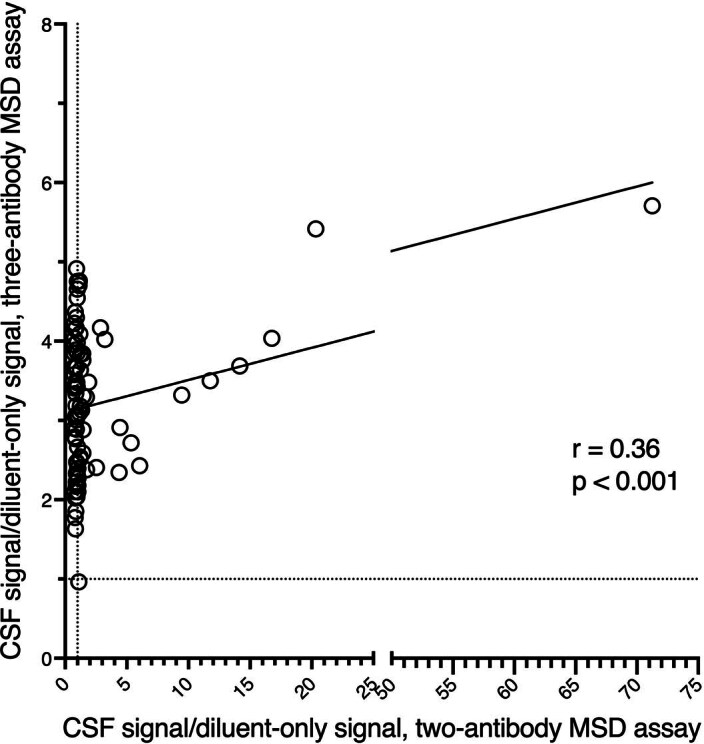
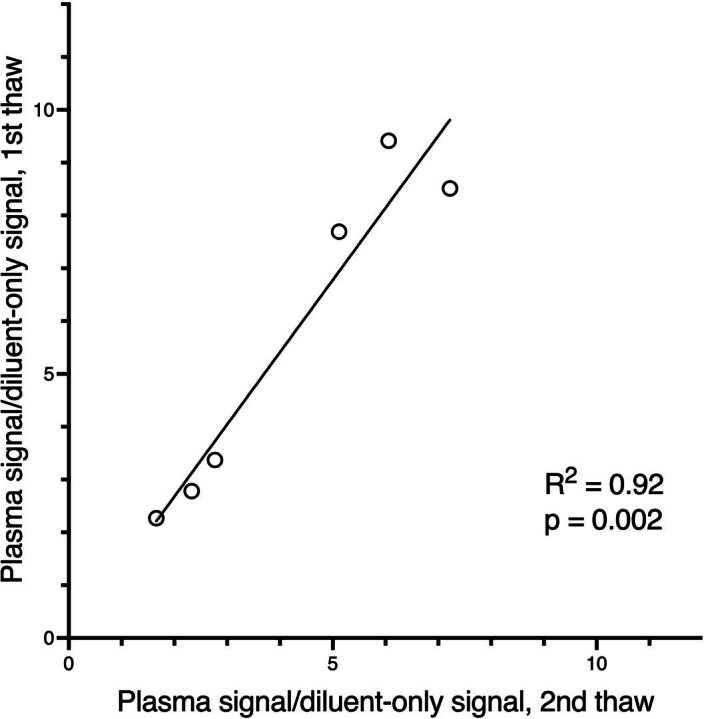
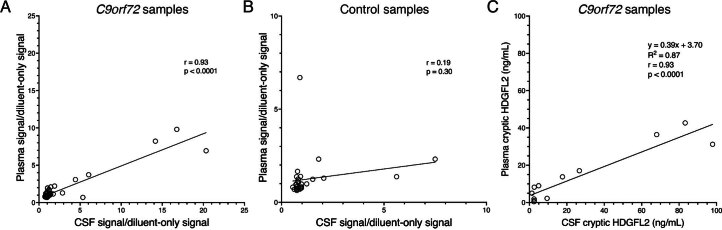
Update of
-
A fluid biomarker reveals loss of TDP-43 splicing repression in pre-symptomatic ALS.bioRxiv [Preprint]. 2023 Jan 24:2023.01.23.525202. doi: 10.1101/2023.01.23.525202. bioRxiv. 2023. Update in: Nat Med. 2024 Feb;30(2):382-393. doi: 10.1038/s41591-023-02788-5. PMID: 36789434 Free PMC article. Updated. Preprint.
References
MeSH terms
Substances
Supplementary concepts
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
