

Bezafibrate attenuates immobilization-induced muscle atrophy in mice
- PMID: 38279013
- PMCID: PMC10817916
- DOI: 10.1038/s41598-024-52689-x
Bezafibrate attenuates immobilization-induced muscle atrophy in mice
Abstract
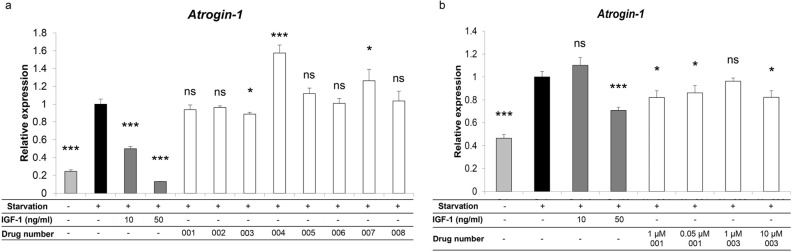
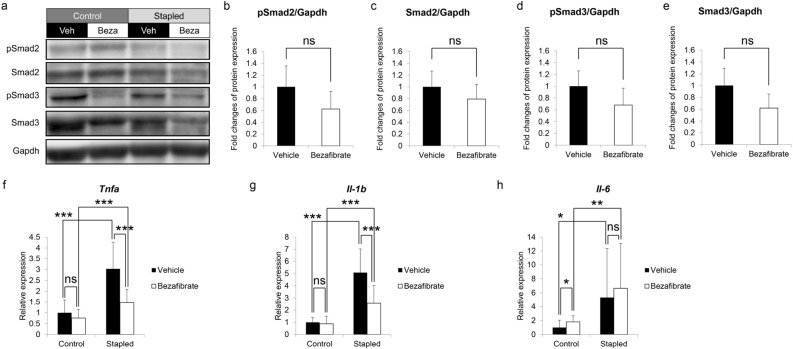
Muscle atrophy due to fragility fractures or frailty worsens not only activity of daily living and healthy life expectancy, but decreases life expectancy. Although several therapeutic agents for muscle atrophy have been investigated, none is yet in clinical use. Here we report that bezafibrate, a drug used to treat hyperlipidemia, can reduce immobilization-induced muscle atrophy in mice. Specifically, we used a drug repositioning approach to screen 144 drugs already utilized clinically for their ability to inhibit serum starvation-induced elevation of Atrogin-1, a factor related to muscle atrophy, in myotubes in vitro. Two candidates were selected, and here we demonstrate that one of them, bezafibrate, significantly reduced muscle atrophy in an in vivo model of muscle atrophy induced by leg immobilization. In gastrocnemius muscle, immobilization reduced muscle weight by an average of ~ 17.2%, and bezafibrate treatment prevented ~ 40.5% of that atrophy. In vitro, bezafibrate significantly inhibited expression of the inflammatory cytokine Tnfa in lipopolysaccharide-stimulated RAW264.7 cells, a murine macrophage line. Finally, we show that expression of Tnfa and IL-1b is induced in gastrocnemius muscle in the leg immobilization model, an activity significantly antagonized by bezafibrate administration in vivo. We conclude that bezafibrate could serve as a therapeutic agent for immobilization-induced muscle atrophy.
© 2024. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures


Similar articles
-
Smad2/3 Proteins Are Required for Immobilization-induced Skeletal Muscle Atrophy.J Biol Chem. 2016 Jun 3;291(23):12184-94. doi: 10.1074/jbc.M115.680579. Epub 2016 Apr 15. J Biol Chem. 2016. PMID: 27129272 Free PMC article.
-
Hemin, heme oxygenase-1 inducer, attenuates immobilization-induced skeletal muscle atrophy in mice.Life Sci. 2013 Apr 9;92(12):740-6. doi: 10.1016/j.lfs.2013.02.008. Epub 2013 Feb 22. Life Sci. 2013. PMID: 23439326
-
[Effects of Atrolnc-1 on immobilization induced muscular atrophy in mice hindlimbs].Zhongguo Ying Yong Sheng Li Xue Za Zhi. 2021 Sep;37(5):566-570. doi: 10.12047/j.cjap.6074.2021.037. Zhongguo Ying Yong Sheng Li Xue Za Zhi. 2021. PMID: 34816675 Chinese.
-
TLR4-defective (C3H/HeJ) mice are not protected from cast immobilization-induced muscle atrophy.Physiol Rep. 2017 Apr;5(8):e13255. doi: 10.14814/phy2.13255. Physiol Rep. 2017. PMID: 28432254 Free PMC article.
-
Flavones Inhibit LPS-Induced Atrogin-1/MAFbx Expression in Mouse C2C12 Skeletal Myotubes.J Nutr Sci Vitaminol (Tokyo). 2015;61(2):188-94. doi: 10.3177/jnsv.61.188. J Nutr Sci Vitaminol (Tokyo). 2015. PMID: 26052151
References
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
