

Mft1, identified from a genome-wide screen of the yeast haploid mutants, mediates cell cycle arrest to counteract quinoxaline-induced toxicity
- PMID: 38283148
- PMCID: PMC10811161
- DOI: 10.3389/fgene.2023.1296383
Mft1, identified from a genome-wide screen of the yeast haploid mutants, mediates cell cycle arrest to counteract quinoxaline-induced toxicity
Abstract
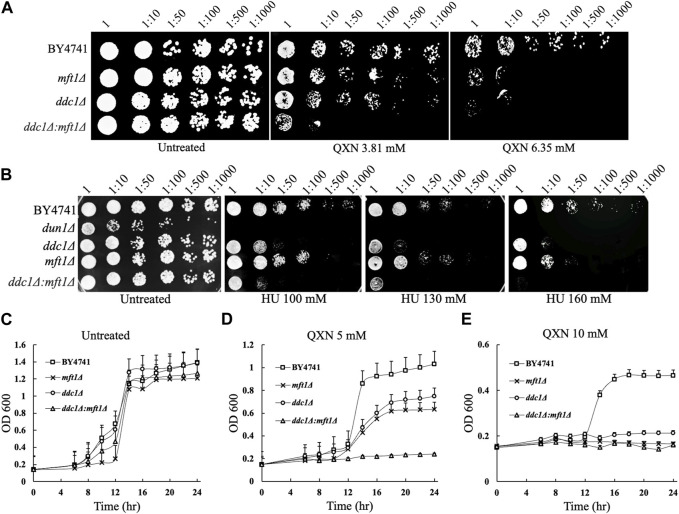
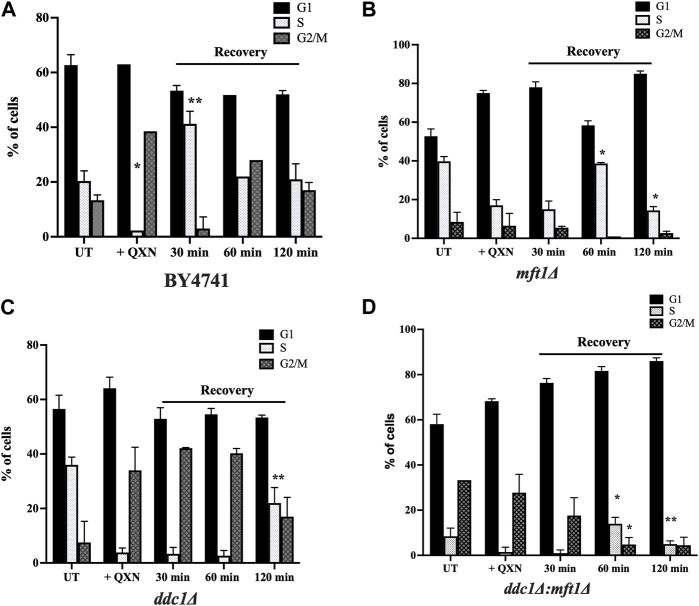
Quinoxaline is a heterocyclic compound with a two-membered ring structure that undergoes redox cycling to produce toxic free radicals. It has antiviral, antibacterial, antifungal, and antitumor activities. However, the biological functions that are involved in mounting a response against the toxic effects of quinoxaline have not been investigated. Herein, we performed a genome-wide screen using the yeast haploid mutant collection and reported the identification of 12 mutants that displayed varying sensitivity towards quinoxaline. No mutant was recovered that showed resistance to quinoxaline. The quinoxaline-sensitive mutants were deleted for genes that encode cell cycle function, as well as genes that belong to other physiological pathways such as the vacuolar detoxification process. Three of the highly sensitive gene-deletion mutants lack the DDC1, DUN1, and MFT1 genes. While Ddc1 and Dun1 are known to perform roles in the cell cycle arrest pathway, the role of Mft1 remains unclear. We show that the mft1Δ mutant is as sensitive to quinoxaline as the ddc1Δ mutant. However, the double mutant ddc1Δ mft1Δ lacking the DDC1 and MFT1 genes, is extremely sensitive to quinoxaline, as compared to the ddc1Δ and mft1Δ single mutants. We further show that the mft1Δ mutant is unable to arrest in the G2/M phase in response to the drug. We conclude that Mft1 performs a unique function independent of Ddc1 in the cell cycle arrest pathway in response to quinoxaline exposure. This is the first demonstration that quinoxaline exerts its toxic effect likely by inducing oxidative DNA damage causing cell cycle arrest. We suggest that clinical applications of quinoxaline and its derivatives should entail targeting cancer cells with defective cell cycle arrest.
Keywords: Saccharomyces cerevisiae; and antitumor activities; antibacterial; antifungal; cell cycle arrest; drug resistance; genome-wide screening; quinoxaline sensitive mutants.
Copyright © 2024 Sulaiman, Al-Ansari, Ali, Aouida and Ramotar.
Conflict of interest statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest. The author(s) declared that they were an editorial board member of Frontiers, at the time of submission. This had no impact on the peer review process and the final decision.
Figures


Similar articles
-
Comparative genome-wide screening identifies a conserved doxorubicin repair network that is diploid specific in Saccharomyces cerevisiae.PLoS One. 2009 Jun 8;4(6):e5830. doi: 10.1371/journal.pone.0005830. PLoS One. 2009. PMID: 19503795 Free PMC article.
-
The novel DNA damage checkpoint protein ddc1p is phosphorylated periodically during the cell cycle and in response to DNA damage in budding yeast.EMBO J. 1997 Sep 1;16(17):5216-26. doi: 10.1093/emboj/16.17.5216. EMBO J. 1997. PMID: 9311982 Free PMC article.
-
DNA-damage induction of RAD54 can be regulated independently of the RAD9- and DDC1-dependent checkpoints that regulate RNR2.Curr Genet. 2002 Jul;41(4):232-40. doi: 10.1007/s00294-002-0302-2. Epub 2002 Jun 27. Curr Genet. 2002. PMID: 12172964
-
Chemical-genetic approaches for exploring the mode of action of natural products.Prog Drug Res. 2008;66:237, 239-71. doi: 10.1007/978-3-7643-8595-8_5. Prog Drug Res. 2008. PMID: 18416308 Review.
-
Dual cell cycle checkpoints sensitive to chromosome replication and DNA damage in the budding yeast Saccharomyces cerevisiae.Radiat Res. 1992 Nov;132(2):141-3. Radiat Res. 1992. PMID: 1438694 Review.
References
-
- Baker Brachmann C., Davies A., Cost G. J., Caputo E., Li J., Hieter P., et al. (1998). Designer deletion strains derived from Saccharomyces cerevisiae S288C: a useful set of strains and plasmids for PCR‐mediated gene disruption and other applications. Yeast 14 (2), 115–132. 10.1002/(SICI)1097-0061(19980130)14:2<115::AID-YEA204>3.0.CO;2-2 - DOI - PubMed
-
- Chandrasekhar S., Reddy N. K., Kumar V. P. (2010). Oxidation of alkynes using PdCl2/CuCl2 in PEG as a recyclable catalytic system: one-pot synthesis of quinoxalines. Tetrahedron Lett. 51 (28), 3623–3625. 10.1016/j.tetlet.2010.05.006 - DOI
-
- Chauhan D. S., Singh P., Quraishi M. (2020). Quinoxaline derivatives as efficient corrosion inhibitors: current status, challenges and future perspectives. J. Mol. Liq. 320, 114387. 10.1016/j.molliq.2020.114387 - DOI
LinkOut - more resources
Full Text Sources