

Dye-Decolorizing Peroxidase of Streptomyces coelicolor (Sc DyPB) Exists as a Dynamic Mixture of Kinetically Different Oligomers
- PMID: 38284010
- PMCID: PMC10809370
- DOI: 10.1021/acsomega.3c07963
Dye-Decolorizing Peroxidase of Streptomyces coelicolor (Sc DyPB) Exists as a Dynamic Mixture of Kinetically Different Oligomers
Abstract
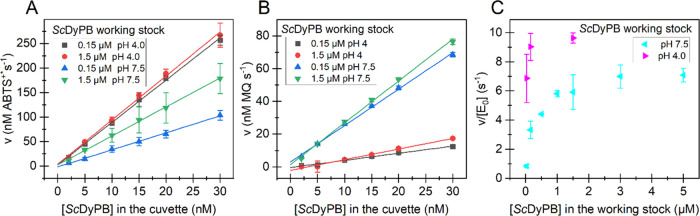
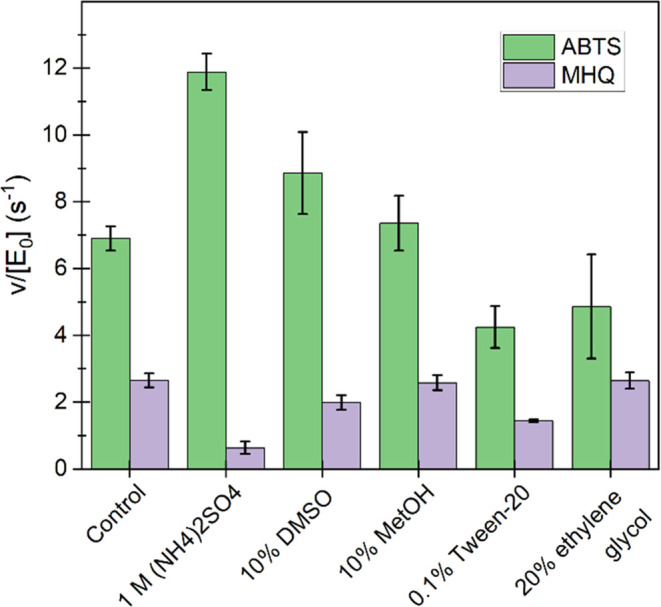
Dye-decolorizing peroxidases (DyPs) are heme-dependent enzymes that catalyze the oxidation of various substrates including environmental pollutants such as azo dyes and also lignin. DyPs often display complex non-Michaelis-Menten kinetics with substrate inhibition or positive cooperativity. Here, we performed in-depth kinetic characterization of the DyP of the bacterium Streptomyces coelicolor (ScDyPB). The activity of ScDyPB was found to be dependent on its concentration in the working stock used to initiate the reactions as well as on the pH of the working stock. Furthermore, the above-listed conditions had different effects on the oxidation of 2,2'-azino-di(3-ethyl-benzothiazoline-6-sulfonic acid) (ABTS) and methylhydroquinone, suggesting that different mechanisms are used in the oxidation of these substrates. The kinetics of the oxidation of ABTS were best described by the model whereby ScDyPB exists as a mixture of two kinetically different enzyme forms. Both forms obey the ping-pong kinetic mechanism, but one form is substrate-inhibited by the ABTS, whereas the other is not. Gel filtration chromatography and dynamic light scattering analyses revealed that ScDyPB exists as a complex mixture of molecules with different sizes. We propose that ScDyPB populations with low and high degrees of oligomerization have different kinetic properties. Such enzyme oligomerization-dependent modulation of the kinetic properties adds further dimension to the complexity of the kinetics of DyPs but also suggests novel possibilities for the regulation of their catalytic activity.
© 2024 The Authors. Published by American Chemical Society.
Conflict of interest statement
The authors declare no competing financial interest.
Figures





References
LinkOut - more resources
Full Text Sources
Other Literature Sources