

Genetically Encoded Photocatalysis Enables Spatially Restricted Optochemical Modulation of Neurons in Live Mice
- PMID: 38292609
- PMCID: PMC10823520
- DOI: 10.1021/acscentsci.3c01351
Genetically Encoded Photocatalysis Enables Spatially Restricted Optochemical Modulation of Neurons in Live Mice
Abstract
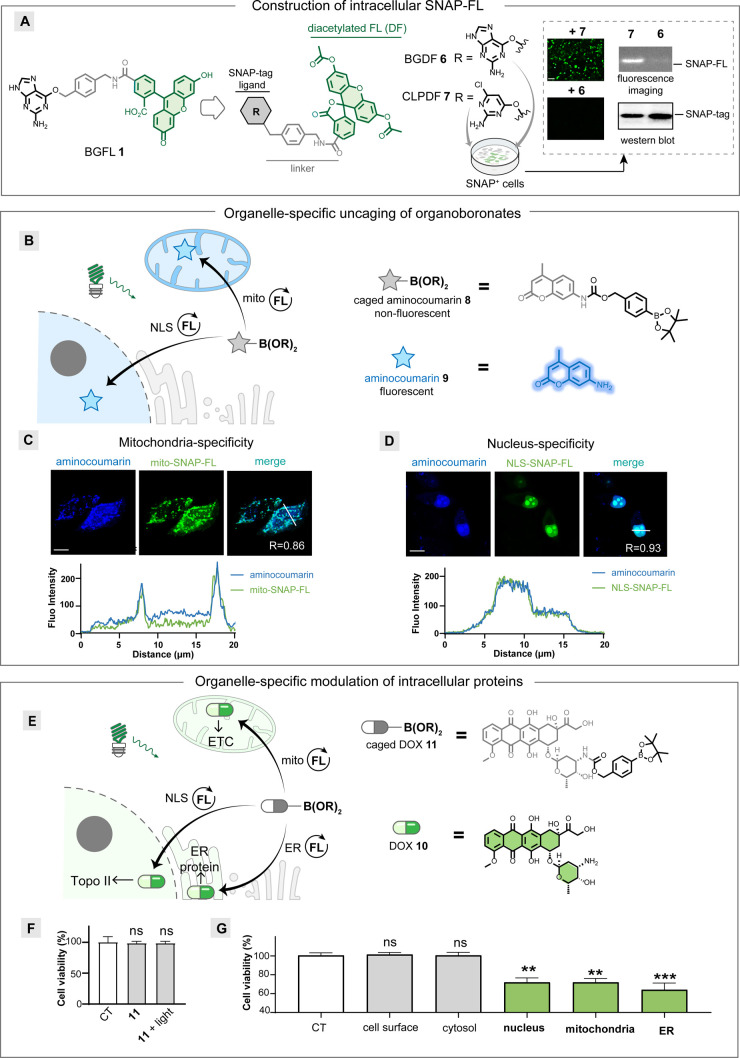
Light provides high temporal precision for neuronal modulations. Small molecules are advantageous for neuronal modulation due to their structural diversity, allowing them to suit versatile targets. However, current optochemical methods release uncaged small molecules with uniform concentrations in the irradiation area, which lack spatial specificity as counterpart optogenetic methods from genetic encoding for photosensitive proteins. Photocatalysis provides spatial specificity by generating reactive species in the proximity of photocatalysts. However, current photocatalytic methods use antibody-tagged heavy-metal photocatalysts for spatial specificity, which are unsuitable for neuronal applications. Here, we report a genetically encoded metal-free photocatalysis method for the optochemical modulation of neurons via deboronative hydroxylation. The genetically encoded photocatalysts generate doxorubicin, a mitochondrial uncoupler, and baclofen by uncaging stable organoboronate precursors. The mitochondria, nucleus, membrane, cytosol, and ER-targeted drug delivery are achieved by this method. The distinct signaling pathway dissection in a single projection is enabled by the dual optogenetic and optochemical control of synaptic transmission. The itching signaling pathway is investigated by photocatalytic uncaging under live-mice skin for the first time by visible light irradiation. The cell-type-specific release of baclofen reveals the GABABR activation on NaV1.8-expressing nociceptor terminals instead of pan peripheral sensory neurons for itch alleviation in live mice.
© 2023 The Authors. Published by American Chemical Society.
Conflict of interest statement
The authors declare no competing financial interest.
Figures





References
-
- Rowland J. M.; van der Plas T. L.; Loidolt M.; Lees R. M.; Keeling J.; Dehning J.; Akam T.; Priesemann V.; Packer A. M. Propagation of activity through the cortical hierarchy and perception are determined by neural variability. Nat. Neurosci. 2023, 26 (9), 1584–1594. 10.1038/s41593-023-01413-5. - DOI - PMC - PubMed
-
- Segal A.; Parkes L.; Aquino K.; Kia S. M.; Wolfers T.; Franke B.; Hoogman M.; Beckmann C. F.; Westlye L. T.; Andreassen O. A.; et al. Regional, circuit and network heterogeneity of brain abnormalities in psychiatric disorders. Nat. Neurosci. 2023, 26 (9), 1613–1629. 10.1038/s41593-023-01404-6. - DOI - PMC - PubMed
LinkOut - more resources
Full Text Sources
Research Materials