

This is a preprint.
Viroid-like colonists of human microbiomes
- PMID: 38293115
- PMCID: PMC10827157
- DOI: 10.1101/2024.01.20.576352
Viroid-like colonists of human microbiomes
Update in
-
Viroid-like colonists of human microbiomes.Cell. 2024 Nov 14;187(23):6521-6536.e18. doi: 10.1016/j.cell.2024.09.033. Epub 2024 Oct 30. Cell. 2024. PMID: 39481381 Free PMC article.
Abstract
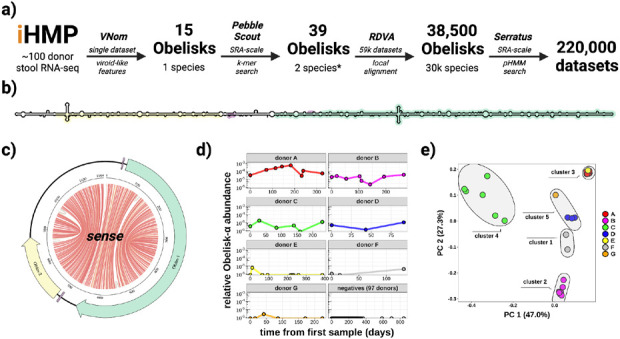
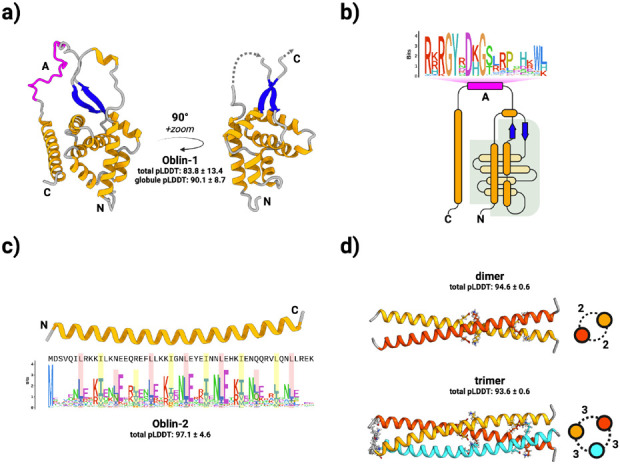
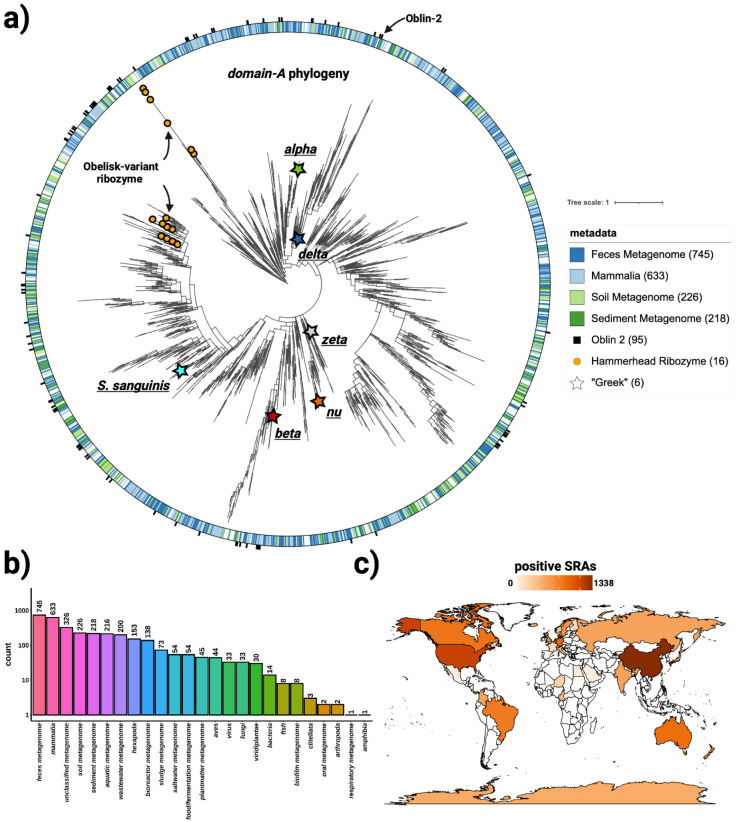
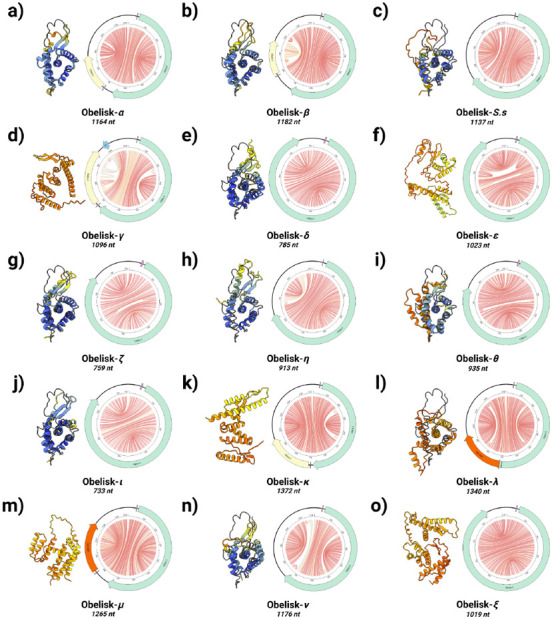
Here, we describe the "Obelisks," a previously unrecognised class of viroid-like elements that we first identified in human gut metatranscriptomic data. "Obelisks" share several properties: (i) apparently circular RNA ~1kb genome assemblies, (ii) predicted rod-like secondary structures encompassing the entire genome, and (iii) open reading frames coding for a novel protein superfamily, which we call the "Oblins". We find that Obelisks form their own distinct phylogenetic group with no detectable sequence or structural similarity to known biological agents. Further, Obelisks are prevalent in tested human microbiome metatranscriptomes with representatives detected in ~7% of analysed stool metatranscriptomes (29/440) and in ~50% of analysed oral metatranscriptomes (17/32). Obelisk compositions appear to differ between the anatomic sites and are capable of persisting in individuals, with continued presence over >300 days observed in one case. Large scale searches identified 29,959 Obelisks (clustered at 90% nucleotide identity), with examples from all seven continents and in diverse ecological niches. From this search, a subset of Obelisks are identified to code for Obelisk-specific variants of the hammerhead type-III self-cleaving ribozyme. Lastly, we identified one case of a bacterial species (Streptococcus sanguinis) in which a subset of defined laboratory strains harboured a specific Obelisk RNA population. As such, Obelisks comprise a class of diverse RNAs that have colonised, and gone unnoticed in, human, and global microbiomes.
Keywords: Hepatitis Delta Virus; Human Microbiome; RNA metaviromics; Viroid.
Figures
References
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous