

Utilization of nanopore direct RNA sequencing to analyze viral RNA modifications
- PMID: 38294229
- PMCID: PMC10878088
- DOI: 10.1128/msystems.01163-23
Utilization of nanopore direct RNA sequencing to analyze viral RNA modifications
Abstract
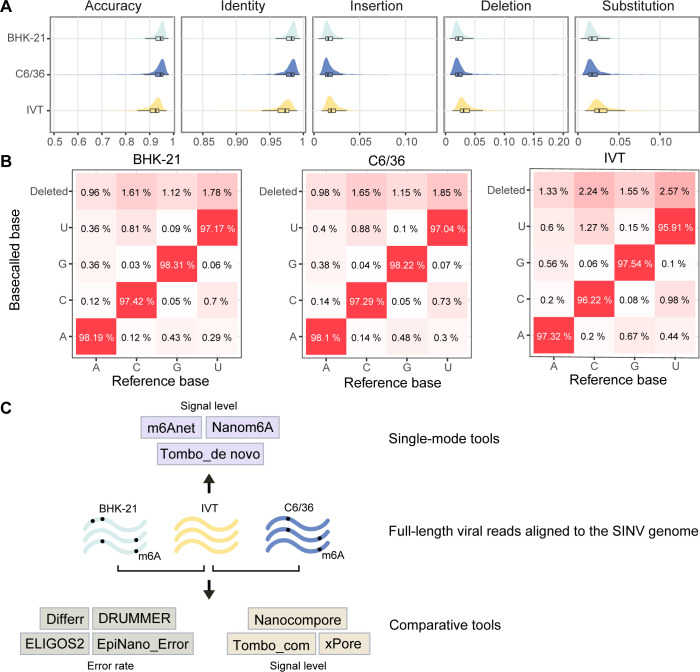
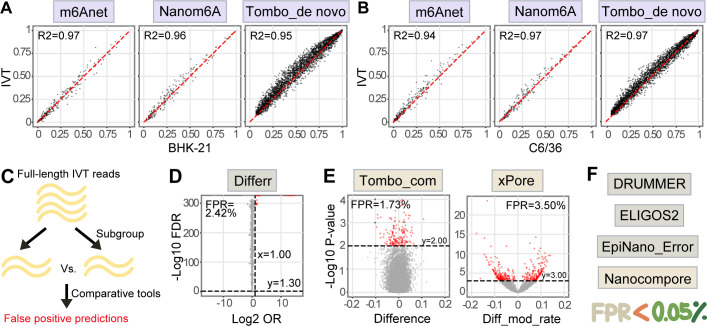
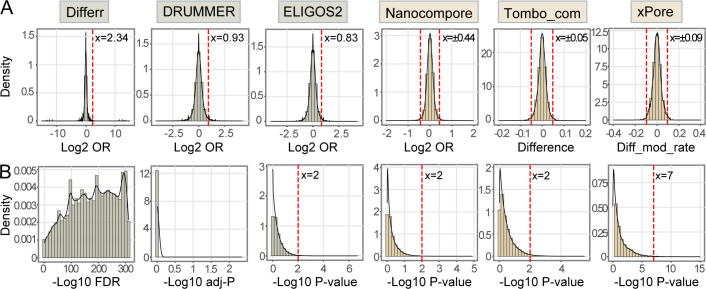
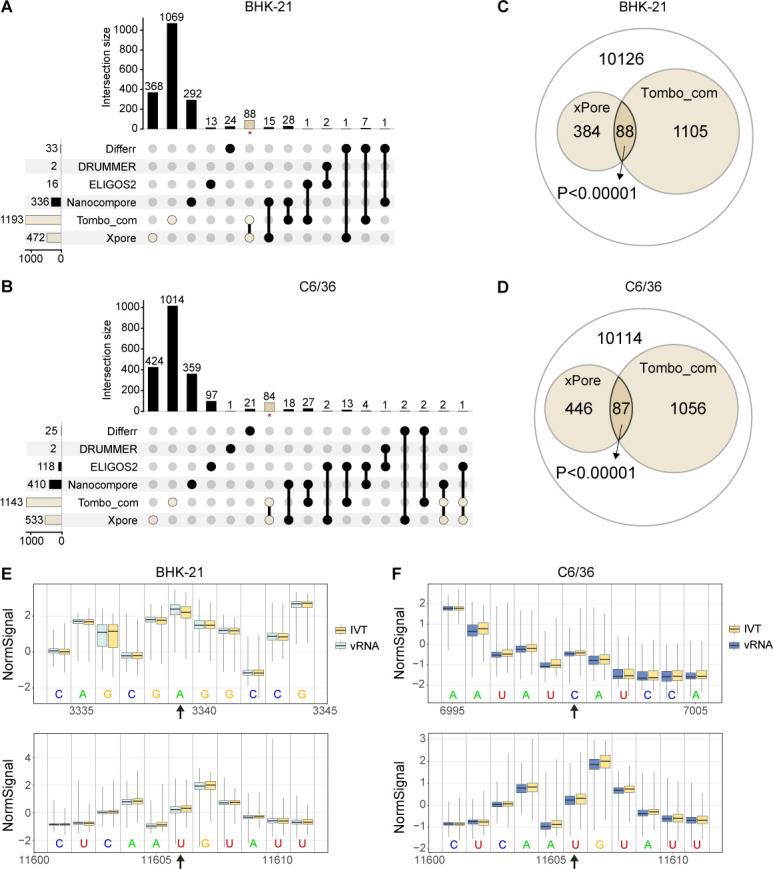
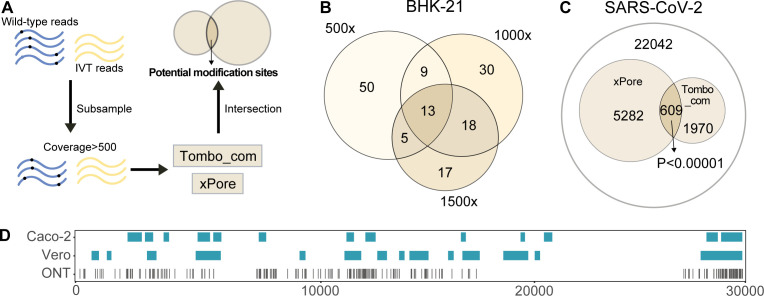
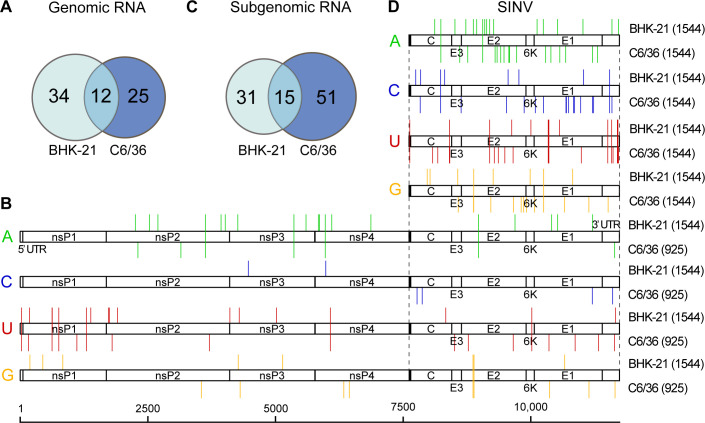
Modifications on viral RNAs (vRNAs), either genomic RNAs or RNA transcripts, have complex effects on the viral life cycle and cellular responses to viral infection. The advent of Oxford Nanopore Technologies Direct RNA Sequencing provides a new strategy for studying RNA modifications. To this end, multiple computational tools have been developed, but a systemic evaluation of their performance in mapping vRNA modifications is lacking. Here, 10 computational tools were tested using the Sindbis virus (SINV) RNAs isolated from infected mammalian (BHK-21) or mosquito (C6/36) cells, with in vitro-transcribed RNAs serving as modification-free control. Three single-mode approaches were shown to be inapplicable in the viral context, and three out of seven comparative methods required cutoff adjustments to reduce false-positive predictions. Utilizing optimized cutoffs, an integrated analysis of comparative tools suggested that the intersected predictions of Tombo_com and xPore were significantly enriched compared with the background. Consequently, a pipeline integrating Tombo_com and xPore was proposed for vRNA modification detection; the performance of which was supported by N6-methyladenosine prediction in severe acute respiratory syndrome coronavirus 2 RNAs using publicly available data. When applied to SINV RNAs, this pipeline revealed more intensive modifications in subgenomic RNAs than in genomic RNAs. Modified uridines were frequently identified, exhibiting substantive overlapping between vRNAs generated in different cell lines. On the other hand, the interpretation of other modifications remained unclear, underlining the limitations of the current computational tools despite their notable potential.IMPORTANCEComputational approaches utilizing Oxford Nanopore Technologies Direct RNA Sequencing data were almost exclusively designed to map eukaryotic epitranscriptomes. Therefore, extra caution must be exercised when using these tools to detect vRNA modifications, as in most cases, vRNA modification profiles should be regarded as unknown epitranscriptomes without prior knowledge. Here, we comprehensively evaluated the performance of 10 computational tools in detecting vRNA modification sites. All tested single-mode methods failed to differentiate native and in vitro-transcribed samples. Using optimized cutoff values, seven tested comparative tools generated very different predictions. An integrated analysis showed significant enrichment of Tombo_com and xPore predictions against the background. A pipeline for vRNA modification detection was proposed accordingly and applied to Sindbis virus RNAs. In conclusion, our study underscores the need for the careful application of computational tools to analyze viral epitranscriptomics. It also offers insights into alphaviral RNA modifications, although further validation is required.
Keywords: RNA modification; Sindbis virus; alphavirus; comparative methods; nanopore.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
Similar articles
-
Identification and Characterization of Sindbis Virus RNA-Host Protein Interactions.J Virol. 2018 Mar 14;92(7):e02171-17. doi: 10.1128/JVI.02171-17. Print 2018 Apr 1. J Virol. 2018. PMID: 29321325 Free PMC article.
-
Treatment of Sindbis Virus-Infected Neurons with Antibody to E2 Alters Synthesis of Complete and nsP1-Expressing Defective Viral RNAs.mBio. 2022 Oct 26;13(5):e0222122. doi: 10.1128/mbio.02221-22. Epub 2022 Sep 7. mBio. 2022. PMID: 36069441 Free PMC article.
-
Analyzing viral epitranscriptomes using nanopore direct RNA sequencing.J Microbiol. 2022 Sep;60(9):867-876. doi: 10.1007/s12275-022-2324-4. Epub 2022 Aug 24. J Microbiol. 2022. PMID: 36001233 Free PMC article.
-
Nanopore-Based Detection of Viral RNA Modifications.mBio. 2022 Jun 28;13(3):e0370221. doi: 10.1128/mbio.03702-21. Epub 2022 May 17. mBio. 2022. PMID: 35579392 Free PMC article. Review.
-
[Advances in mapping analysis of ribonucleic acid modifications through sequencing].Se Pu. 2024 Jul;42(7):632-645. doi: 10.3724/SP.J.1123.2023.12025. Se Pu. 2024. PMID: 38966972 Free PMC article. Review. Chinese.
Cited by
-
Wolbachia elevates host methyltransferase expression and alters the m6A methylation landscape in Aedes aegypti mosquito cells.BMC Microbiol. 2025 Mar 25;25(1):164. doi: 10.1186/s12866-025-03898-5. BMC Microbiol. 2025. PMID: 40128692 Free PMC article.
-
Analysis of bacterial transcriptome and epitranscriptome using nanopore direct RNA sequencing.Nucleic Acids Res. 2024 Aug 27;52(15):8746-8762. doi: 10.1093/nar/gkae601. Nucleic Acids Res. 2024. PMID: 39011882 Free PMC article.
References
-
- Boccaletto P, Stefaniak F, Ray A, Cappannini A, Mukherjee S, Purta E, Kurkowska M, Shirvanizadeh N, Destefanis E, Groza P, Avşar G, Romitelli A, Pir P, Dassi E, Conticello SG, Aguilo F, Bujnicki JM. 2022. MODOMICS: a database of RNA modification pathways. 2021 update. Nucleic Acids Res 50:D231–D235. doi:10.1093/nar/gkab1083 - DOI - PMC - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
