

A synthetic biology approach to assemble and reboot clinically relevant Pseudomonas aeruginosa tailed phages
- PMID: 38294230
- PMCID: PMC10913387
- DOI: 10.1128/spectrum.02897-23
A synthetic biology approach to assemble and reboot clinically relevant Pseudomonas aeruginosa tailed phages
Abstract
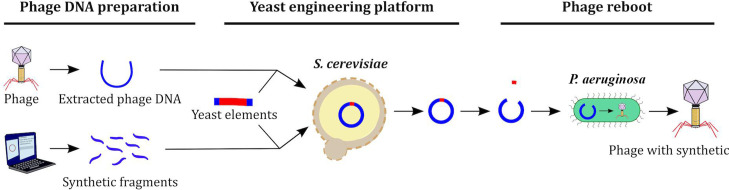
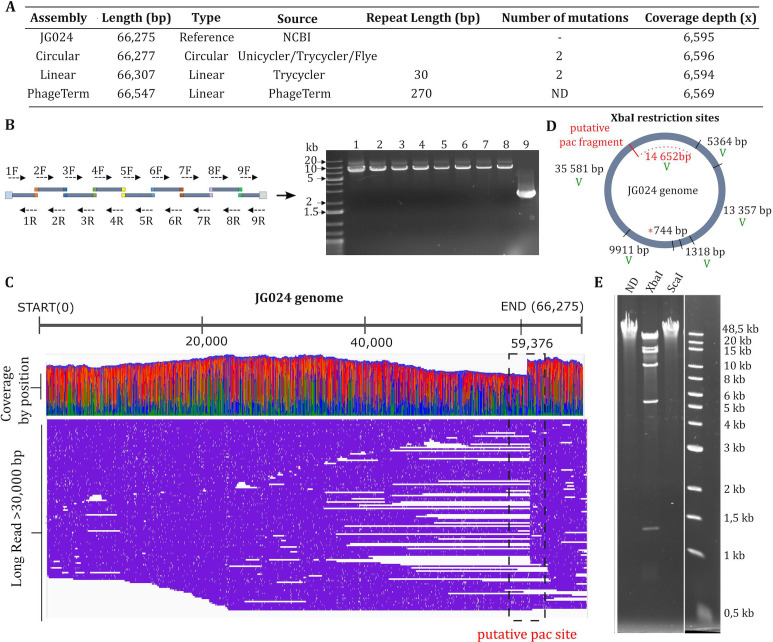
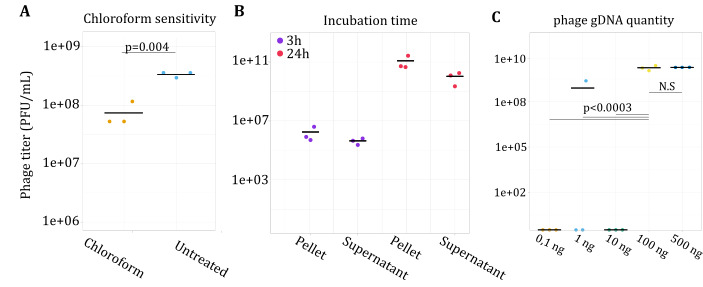
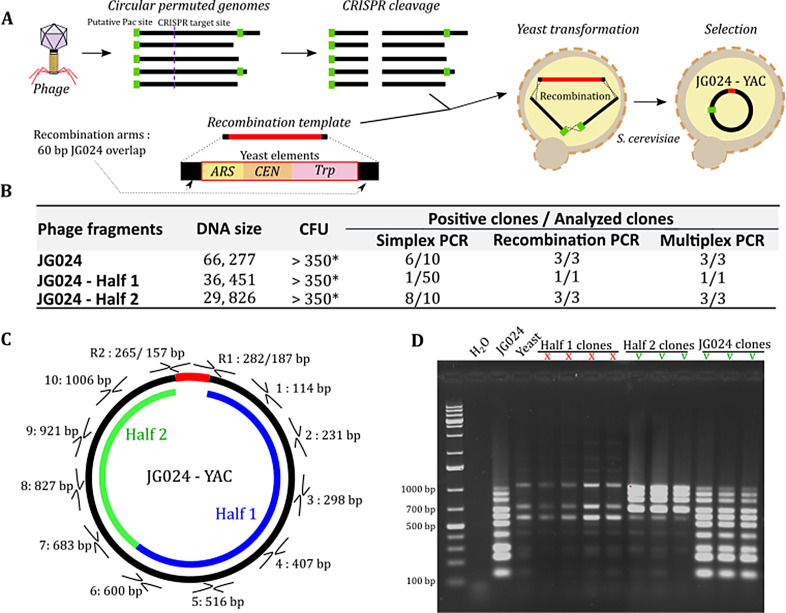
The rise in the frequency of antibiotic resistance has made bacterial infections, specifically Pseudomonas aeruginosa, a cause for greater concern. Phage therapy is a promising solution that uses naturally isolated phages to treat bacterial infections. Ecological limitations, which stipulate a discrete host range and the inevitable evolution of resistance, may be overcome through a better understanding of phage biology and the utilization of engineered phages. In this study, we developed a synthetic biology approach to construct tailed phages that naturally target clinically relevant strains of Pseudomonas aeruginosa. As proof of concept, we successfully cloned and assembled the JG024 and DMS3 phage genomes in yeast using transformation-associated recombination cloning and rebooted these two phage genomes in two different strains of P. aeruginosa. We identified factors that affected phage reboot efficiency like the phage species or the presence of antiviral defense systems in the bacterial strain. We have successfully extended this method to two other phage species and observed that the method enables the reboot of phages that are naturally unable to infect the strain used for reboot. This research represents a critical step toward the construction of clinically relevant, engineered P. aeruginosa phages.IMPORTANCEPseudomonas aeruginosa is a bacterium responsible for severe infections and a common major complication in cystic fibrosis. The use of antibiotics to treat bacterial infections has become increasingly difficult as antibiotic resistance has become more prevalent. Phage therapy is an alternative solution that is already being used in some European countries, but its use is limited by the narrow host range due to the phage receptor specificity, the presence of antiviral defense systems in the bacterial strain, and the possible emergence of phage resistance. In this study, we demonstrate the use of a synthetic biology approach to construct and reboot clinically relevant P. aeruginosa tailed phages. This method enables a significant expansion of possibilities through the construction of engineered phages for therapy applications.
Keywords: Pseudomonas aeruginosa; phage reboot; phage therapy; synthetic biology.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
Similar articles
-
Characterization of a new Pseudomonas aeruginosa Queuovirinae bacteriophage.Microbiol Spectr. 2024 Mar 5;12(3):e0371923. doi: 10.1128/spectrum.03719-23. Epub 2024 Feb 12. Microbiol Spectr. 2024. PMID: 38345389 Free PMC article.
-
Isolation and Characterization of Three Pseudomonas aeruginosa Viruses with Therapeutic Potential.Microbiol Spectr. 2023 Jun 15;11(3):e0463622. doi: 10.1128/spectrum.04636-22. Epub 2023 May 1. Microbiol Spectr. 2023. PMID: 37125933 Free PMC article.
-
Study of the probability of resistance to phage infection in a collection of clinical isolates of Pseudomonas aeruginosa in relation to the presence of Pf phages.Microbiol Spectr. 2025 Mar 4;13(3):e0301024. doi: 10.1128/spectrum.03010-24. Epub 2025 Feb 5. Microbiol Spectr. 2025. PMID: 39907445 Free PMC article.
-
Pseudomonas aeruginosa Bacteriophages and Their Clinical Applications.Viruses. 2024 Jun 29;16(7):1051. doi: 10.3390/v16071051. Viruses. 2024. PMID: 39066214 Free PMC article. Review.
-
Bacteriophages of Pseudomonas aeruginosa: long-term prospects for use in phage therapy.Adv Virus Res. 2014;88:227-78. doi: 10.1016/B978-0-12-800098-4.00005-2. Adv Virus Res. 2014. PMID: 24373314 Review.
Cited by
-
Synthetic and Functional Engineering of Bacteriophages: Approaches for Tailored Bactericidal, Diagnostic, and Delivery Platforms.Molecules. 2025 Jul 25;30(15):3132. doi: 10.3390/molecules30153132. Molecules. 2025. PMID: 40807307 Free PMC article. Review.
-
Phage Therapy: An Alternative Approach to Combating Multidrug-Resistant Bacterial Infections in Cystic Fibrosis.Int J Mol Sci. 2024 Jul 30;25(15):8321. doi: 10.3390/ijms25158321. Int J Mol Sci. 2024. PMID: 39125890 Free PMC article. Review.
-
Protocol for end-design-free rebooting of terminally redundant Pseudomonas phages in clinical isolates of Pseudomonas aeruginosa.STAR Protoc. 2025 Aug 4;6(3):104012. doi: 10.1016/j.xpro.2025.104012. Online ahead of print. STAR Protoc. 2025. PMID: 40763034 Free PMC article.
-
Replacement of the Genomic Scaffold Improves the Replication Efficiency of Synthetic Klebsiella Phages.Int J Mol Sci. 2025 Jul 16;26(14):6824. doi: 10.3390/ijms26146824. Int J Mol Sci. 2025. PMID: 40725069 Free PMC article.
References
-
- ECSF . 2020. European cystic fibrosis society patient registry annual report 2020. Available from: https://www.ecfs.eu/sites/default/files/ECFSPR_Report_2020_v1.0%2807Jun2...
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Research Materials
