

How intrinsically disordered proteins order plant gene silencing
- PMID: 38296708
- PMCID: PMC10932933
- DOI: 10.1016/j.tig.2023.12.009
How intrinsically disordered proteins order plant gene silencing
Abstract
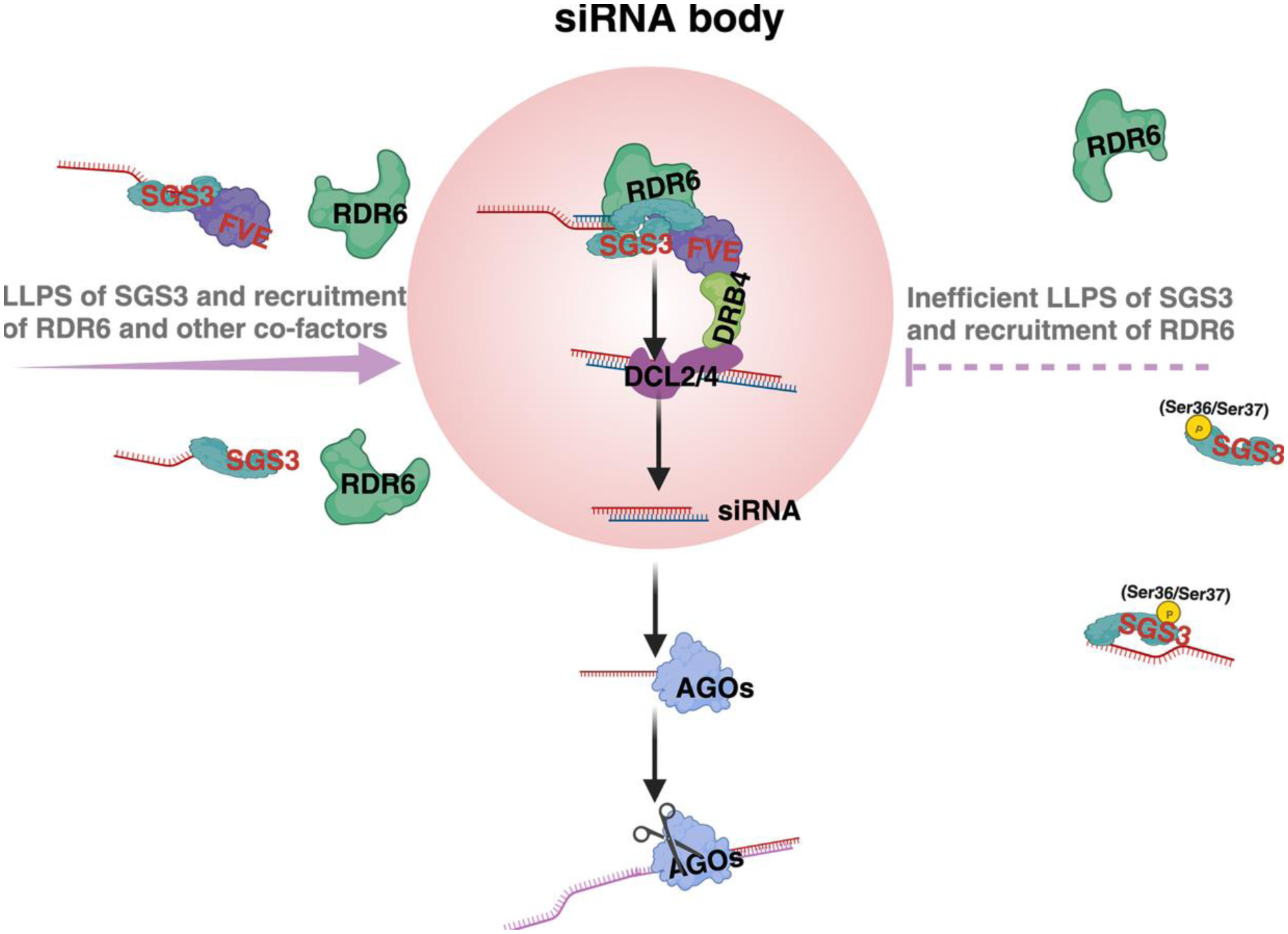
Intrinsically disordered proteins (IDPs) and proteins with intrinsically disordered regions (IDRs) possess low sequence complexity of amino acids and display non-globular tertiary structures. They can act as scaffolds, form regulatory hubs, or trigger biomolecular condensation to control diverse aspects of biology. Emerging evidence has recently implicated critical roles of IDPs and IDR-contained proteins in nuclear transcription and cytoplasmic post-transcriptional processes, among other molecular functions. We here summarize the concepts and organizing principles of IDPs. We then illustrate recent progress in understanding the roles of key IDPs in machineries that regulate transcriptional and post-transcriptional gene silencing (PTGS) in plants, aiming at highlighting new modes of action of IDPs in controlling biological processes.
Keywords: gene silencing; intrinsically disordered protein (IDP); intrinsically disordered region (IDR); liquid–liquid phase separation; miRNA; siRNA.
Published by Elsevier Ltd.
Conflict of interest statement
Declaration of interests The authors declare no conflicts of interests.
Figures



References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
