

METTL16 promotes liver cancer stem cell self-renewal via controlling ribosome biogenesis and mRNA translation
- PMID: 38302992
- PMCID: PMC10835888
- DOI: 10.1186/s13045-024-01526-9
METTL16 promotes liver cancer stem cell self-renewal via controlling ribosome biogenesis and mRNA translation
Abstract
Background: While liver cancer stem cells (CSCs) play a crucial role in hepatocellular carcinoma (HCC) initiation, progression, recurrence, and treatment resistance, the mechanism underlying liver CSC self-renewal remains elusive. We aim to characterize the role of Methyltransferase 16 (METTL16), a recently identified RNA N6-methyladenosine (m6A) methyltransferase, in HCC development/maintenance, CSC stemness, as well as normal hepatogenesis.
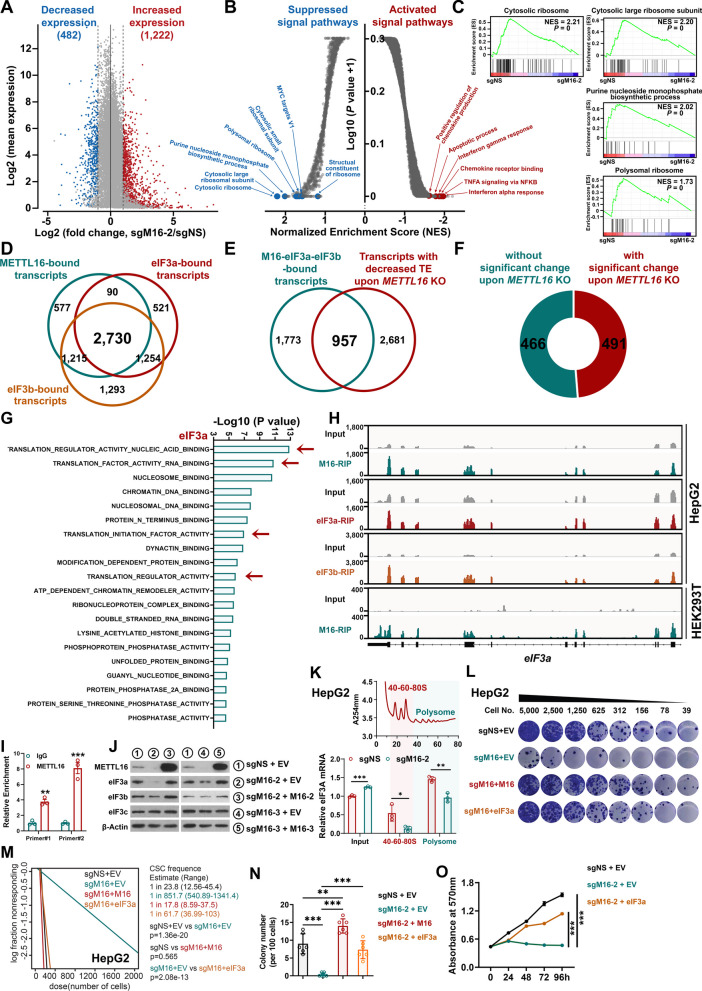
Methods: Liver-specific Mettl16 conditional KO (cKO) mice were generated to assess its role in HCC pathogenesis and normal hepatogenesis. Hydrodynamic tail-vein injection (HDTVi)-induced de novo hepatocarcinogenesis and xenograft models were utilized to determine the role of METTL16 in HCC initiation and progression. A limiting dilution assay was utilized to evaluate CSC frequency. Functionally essential targets were revealed via integrative analysis of multi-omics data, including RNA-seq, RNA immunoprecipitation (RIP)-seq, and ribosome profiling.
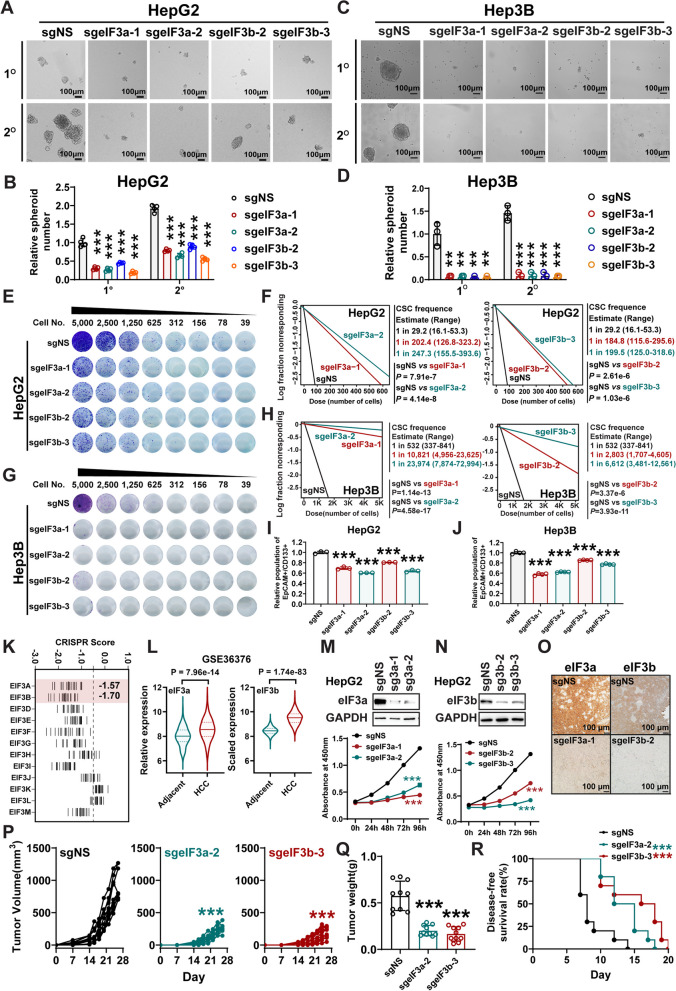
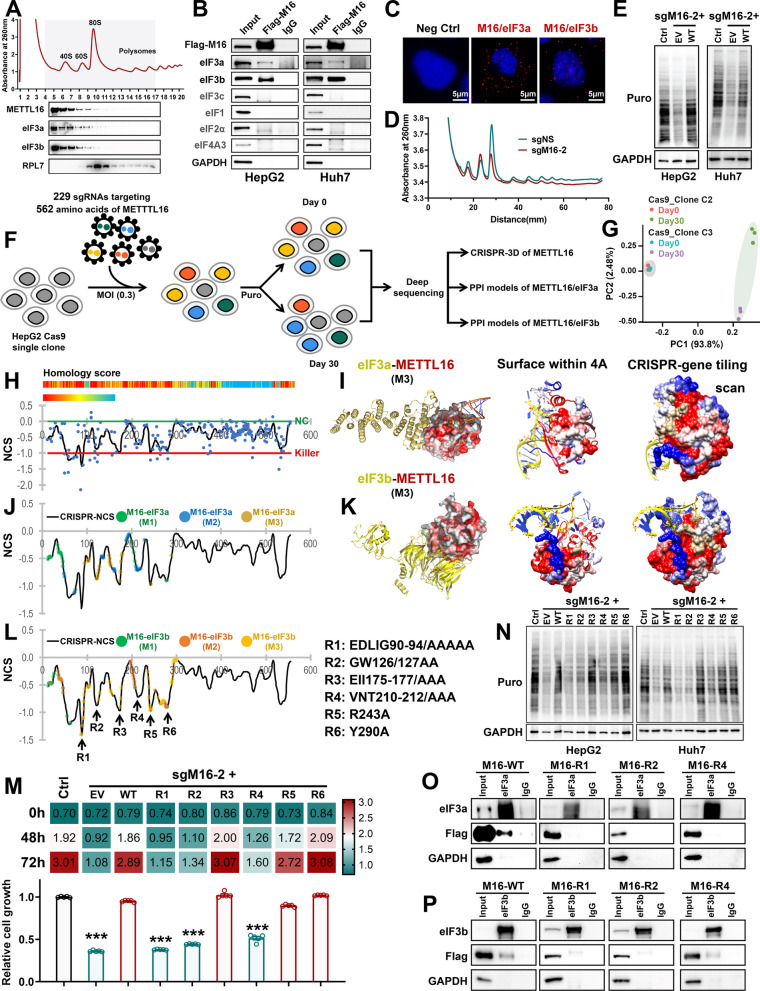
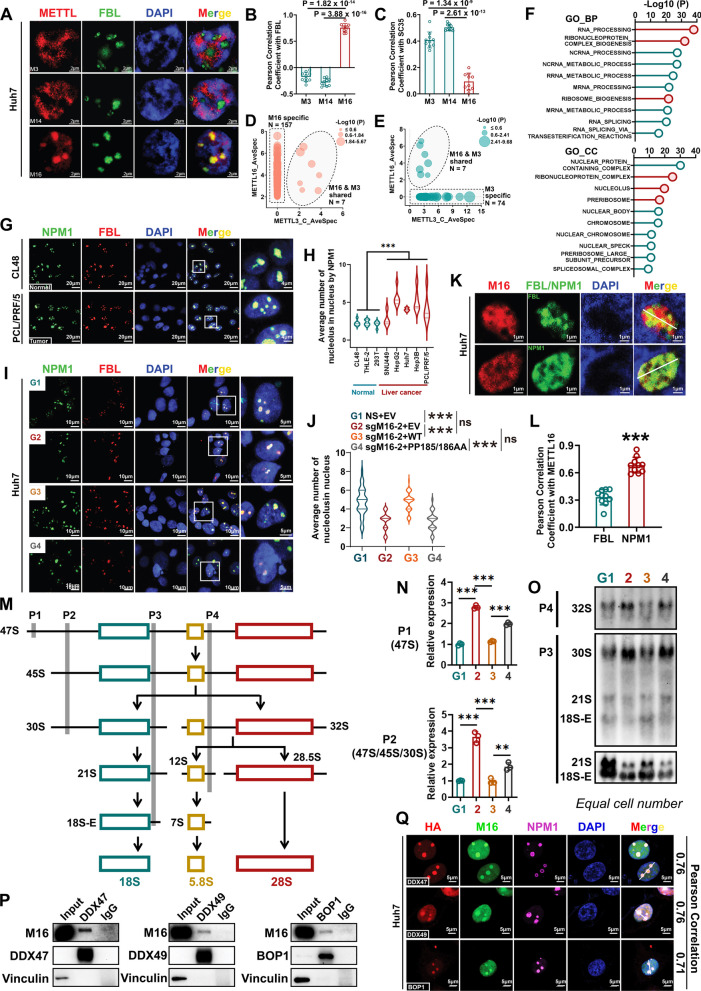
Results: METTL16 is highly expressed in liver CSCs and its depletion dramatically decreased CSC frequency in vitro and in vivo. Mettl16 KO significantly attenuated HCC initiation and progression, yet only slightly influenced normal hepatogenesis. Mechanistic studies, including high-throughput sequencing, unveiled METTL16 as a key regulator of ribosomal RNA (rRNA) maturation and mRNA translation and identified eukaryotic translation initiation factor 3 subunit a (eIF3a) transcript as a bona-fide target of METTL16 in HCC. In addition, the functionally essential regions of METTL16 were revealed by CRISPR gene tiling scan, which will pave the way for the development of potential inhibitor(s).
Conclusions: Our findings highlight the crucial oncogenic role of METTL16 in promoting HCC pathogenesis and enhancing liver CSC self-renewal through augmenting mRNA translation efficiency.
Keywords: Cancer stem cells; Hepatocellular carcinoma; METTL16; N6-methyladenosine; Ribosome biogenesis; Self-renewal; eIF3a; mRNA translation.
© 2024. The Author(s).
Conflict of interest statement
All authors declare no potential conflicts of interest.
Figures


References
-
- Yu H, Zhou L, Loong JHC, et al. SERPINA12 promotes the tumorigenic capacity of HCC stem cells through hyperactivation of AKT/beta-catenin signaling. Hepatology. 2023;24:9173. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
