

Extracellular matrix protein composition dynamically changes during murine forelimb development
- PMID: 38303699
- PMCID: PMC10831947
- DOI: 10.1016/j.isci.2024.108838
Extracellular matrix protein composition dynamically changes during murine forelimb development
Abstract
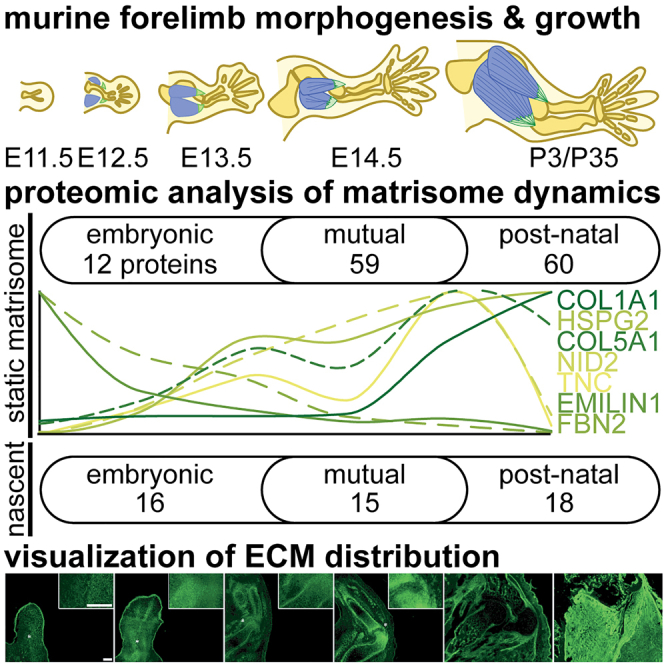
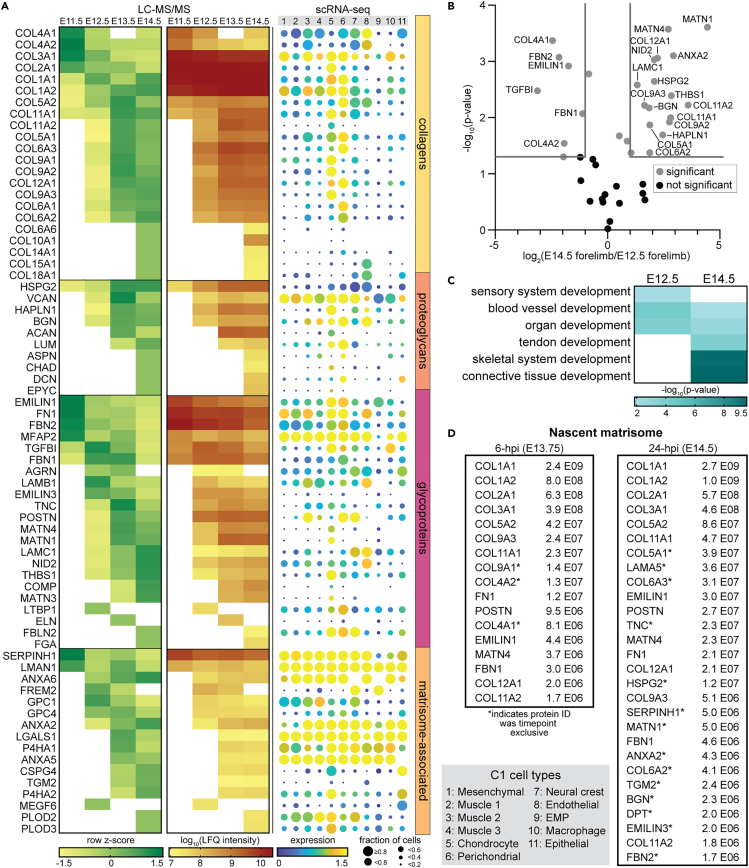
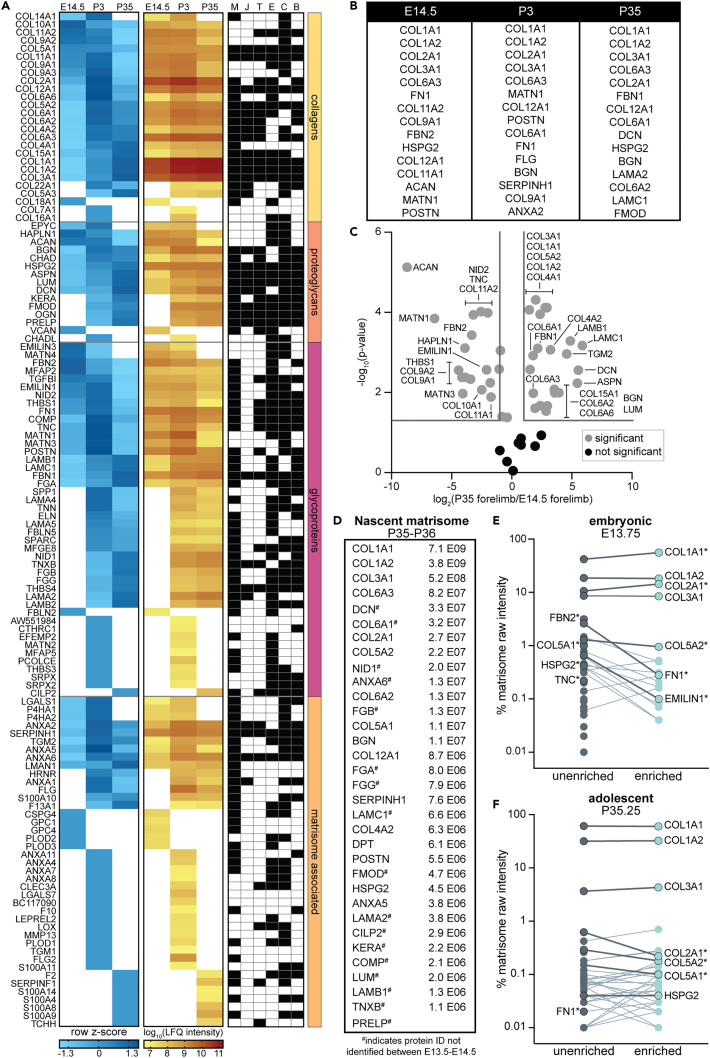
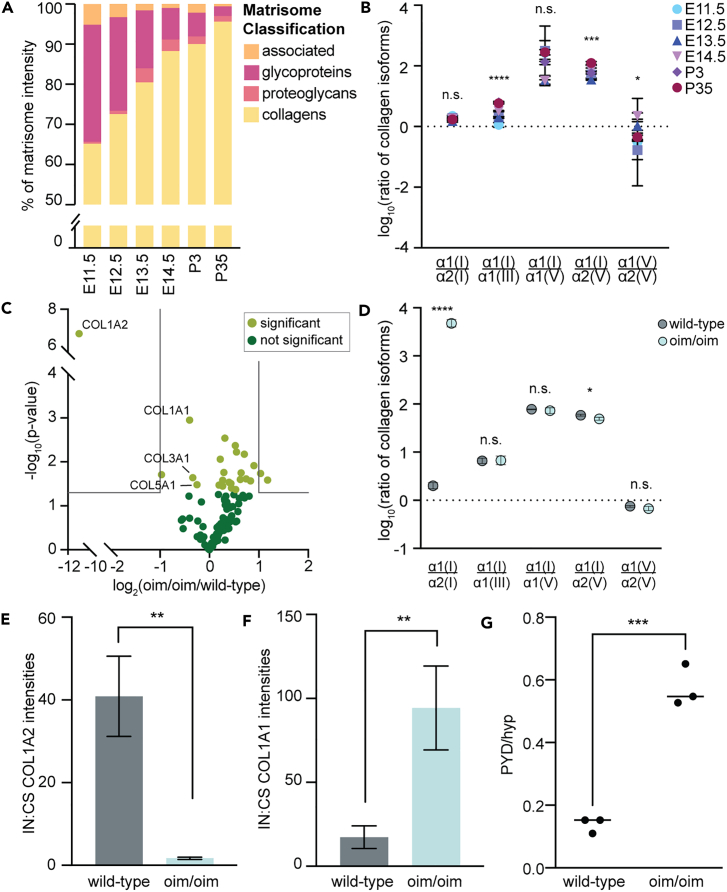
The extracellular matrix (ECM) is an integral part of multicellular organisms, connecting different cell layers and tissue types. During morphogenesis and growth, tissues undergo substantial reorganization. While it is intuitive that the ECM remodels in concert, little is known regarding how matrix composition and organization change during development. Here, we quantified ECM protein dynamics in the murine forelimb during appendicular musculoskeletal morphogenesis (embryonic days 11.5-14.5) using tissue fractionation, bioorthogonal non-canonical amino acid tagging, and mass spectrometry. Our analyses indicated that ECM protein (matrisome) composition in the embryonic forelimb changed as a function of development and growth, was distinct from other developing organs (brain), and was altered in a model of disease (osteogenesis imperfecta murine). Additionally, the tissue distribution for select matrisome was assessed via immunohistochemistry in the wild-type embryonic and postnatal musculoskeletal system. This resource will guide future research investigating the role of the matrisome during complex tissue development.
Keywords: Biological sciences; Developmental biology; Natural sciences.
© 2024 The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures


References
-
- McQueen C., Towers M. Establishing the pattern of the vertebrate limb. Development. 2020;147 - PubMed
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
