

Targeting circulating labile heme as a defense strategy against malaria
- PMID: 38307624
- PMCID: PMC10837040
- DOI: 10.26508/lsa.202302276
Targeting circulating labile heme as a defense strategy against malaria
Abstract
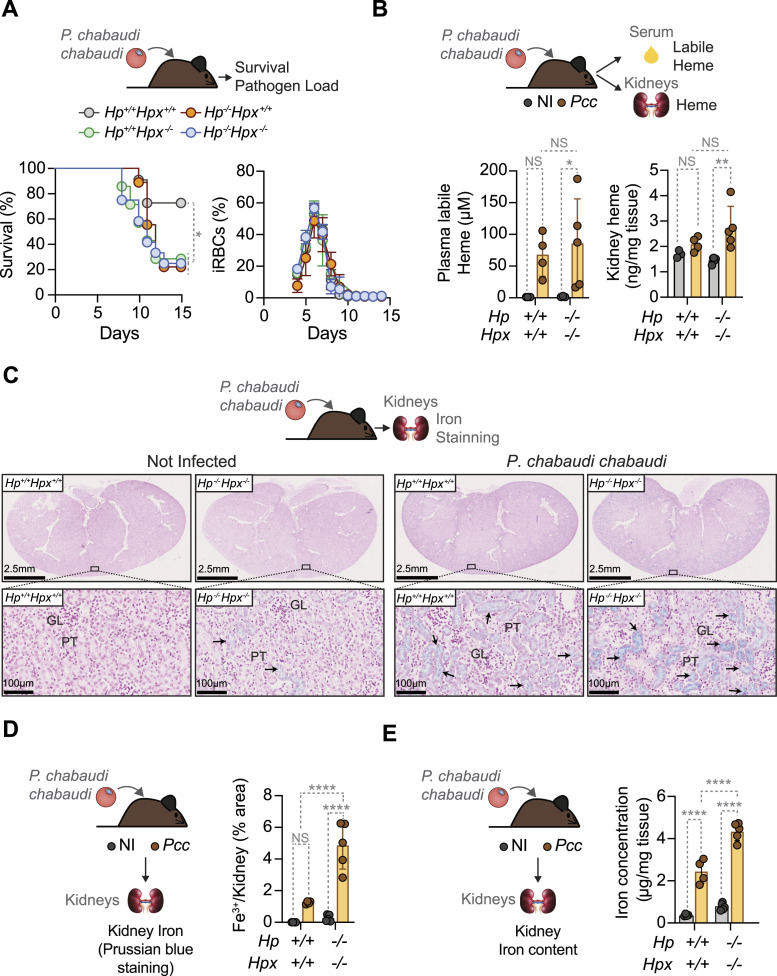
Severe presentations of malaria emerge as Plasmodium (P.) spp. parasites invade and lyse red blood cells (RBC), producing extracellular hemoglobin (HB), from which labile heme is released. Here, we tested whether scavenging of extracellular HB and/or labile heme, by haptoglobin (HP) and/or hemopexin (HPX), respectively, counter the pathogenesis of severe presentations of malaria. We found that circulating labile heme is an independent risk factor for cerebral and non-cerebral presentations of severe P. falciparum malaria in children. Labile heme was negatively correlated with circulating HP and HPX, which were, however, not risk factors for severe P. falciparum malaria. Genetic Hp and/or Hpx deletion in mice led to labile heme accumulation in plasma and kidneys, upon Plasmodium infection This was associated with higher incidence of mortality and acute kidney injury (AKI) in ageing but not adult Plasmodium-infected mice, and was corroborated by an inverse correlation between heme and HPX with serological markers of AKI in P. falciparum malaria. In conclusion, HP and HPX act in an age-dependent manner to prevent the pathogenesis of severe presentation of malaria in mice and presumably in humans.
© 2024 Ramos et al.
Conflict of interest statement
The authors declare that they have no conflict of interest.
Figures








References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous