

Repurposing of Zika virus live-attenuated vaccine (ZIKV-LAV) strains as oncolytic viruses targeting human glioblastoma multiforme cells
- PMID: 38308299
- PMCID: PMC10835997
- DOI: 10.1186/s12967-024-04930-4
Repurposing of Zika virus live-attenuated vaccine (ZIKV-LAV) strains as oncolytic viruses targeting human glioblastoma multiforme cells
Abstract
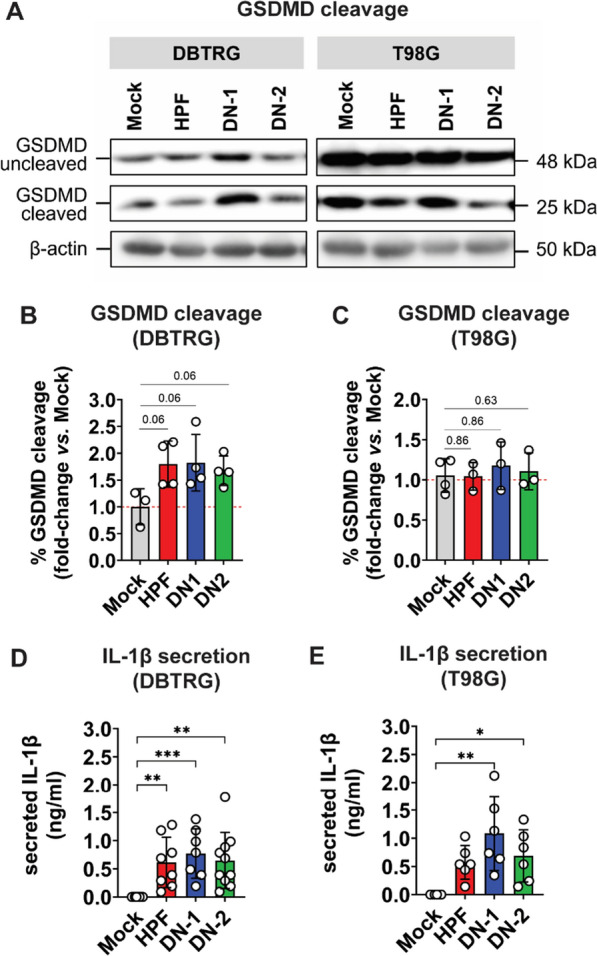
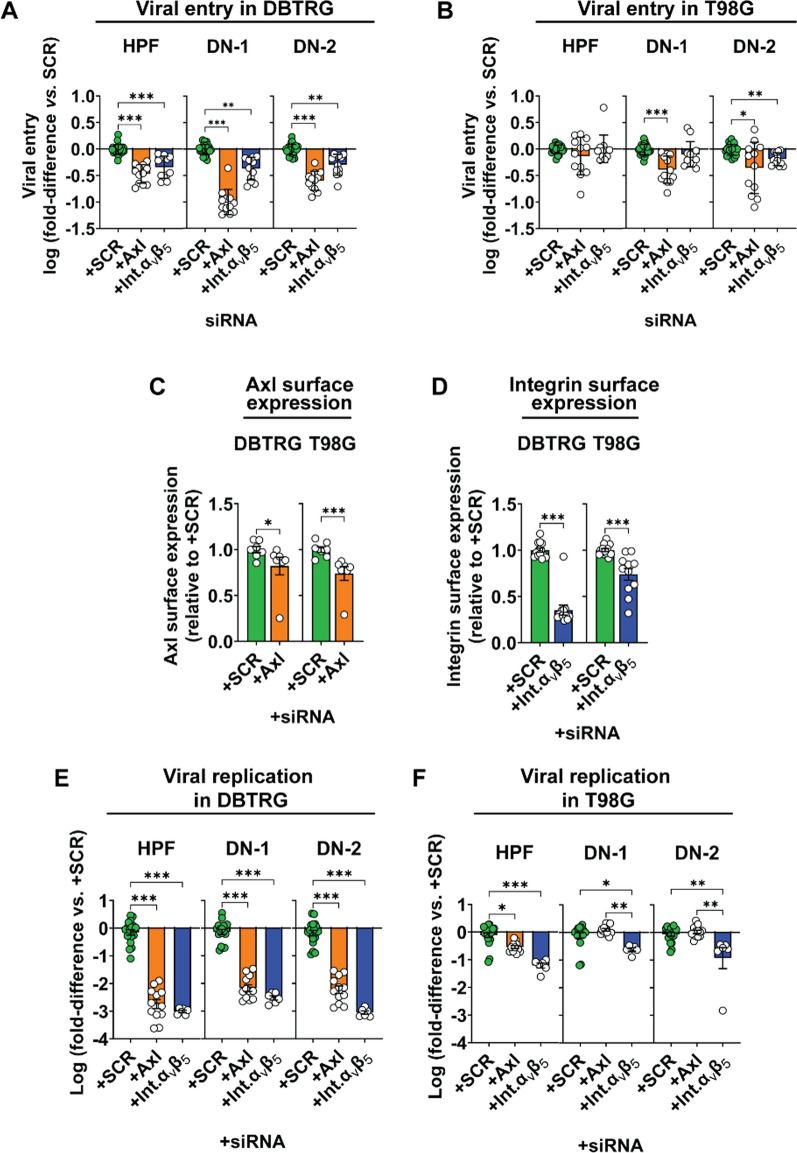
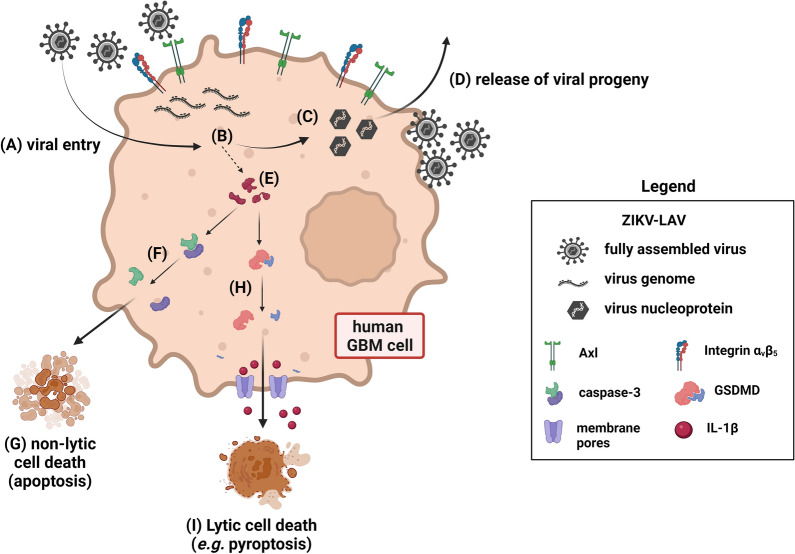
Glioblastoma multiforme (GBM) is the most common malignant primary brain cancer affecting the adult population. Median overall survival for GBM patients is poor (15 months), primarily due to high rates of tumour recurrence and the paucity of treatment options. Oncolytic virotherapy is a promising treatment alternative for GBM patients, where engineered viruses selectively infect and eradicate cancer cells by inducing cell lysis and eliciting robust anti-tumour immune response. In this study, we evaluated the oncolytic potency of live-attenuated vaccine strains of Zika virus (ZIKV-LAV) against human GBM cells in vitro. Our findings revealed that Axl and integrin αvβ5 function as cellular receptors mediating ZIKV-LAV infection in GBM cells. ZIKV-LAV strains productively infected and lysed human GBM cells but not primary endothelia and terminally differentiated neurons. Upon infection, ZIKV-LAV mediated GBM cell death via apoptosis and pyroptosis. This is the first in-depth molecular dissection of how oncolytic ZIKV infects and induces death in tumour cells.
Keywords: GBM; Glioblastoma; Immunogenic cell death; Live-attenuated vaccine; Oncolytic virus; ZIKV; Zika virus.
© 2024. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures
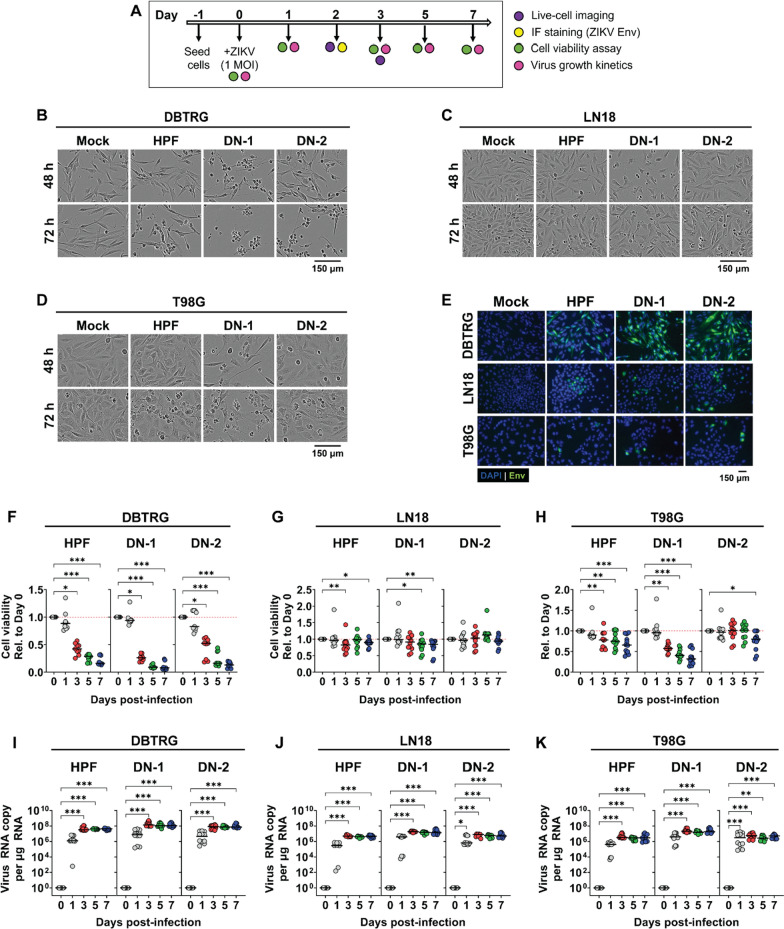
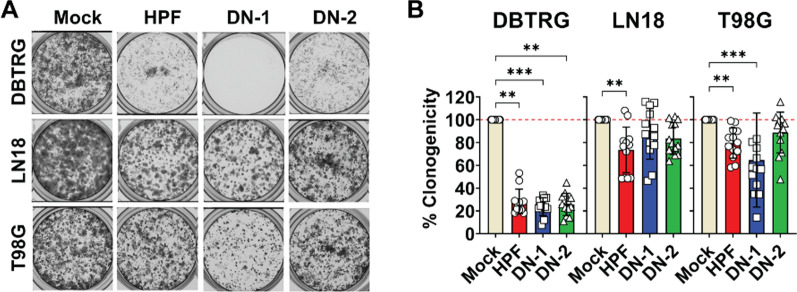
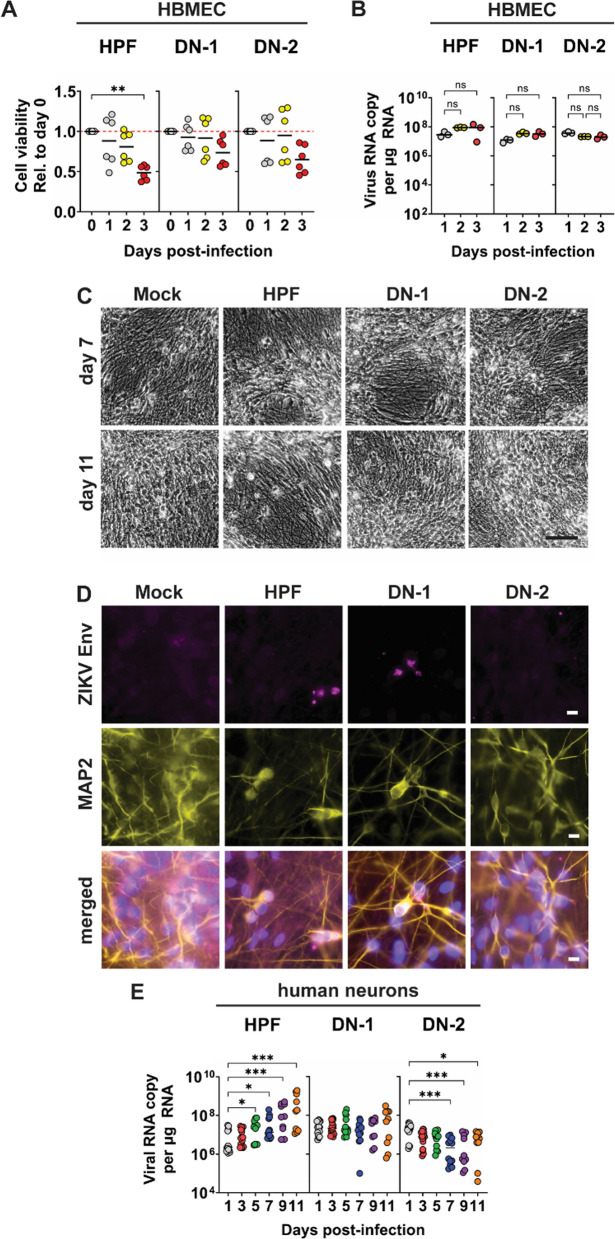
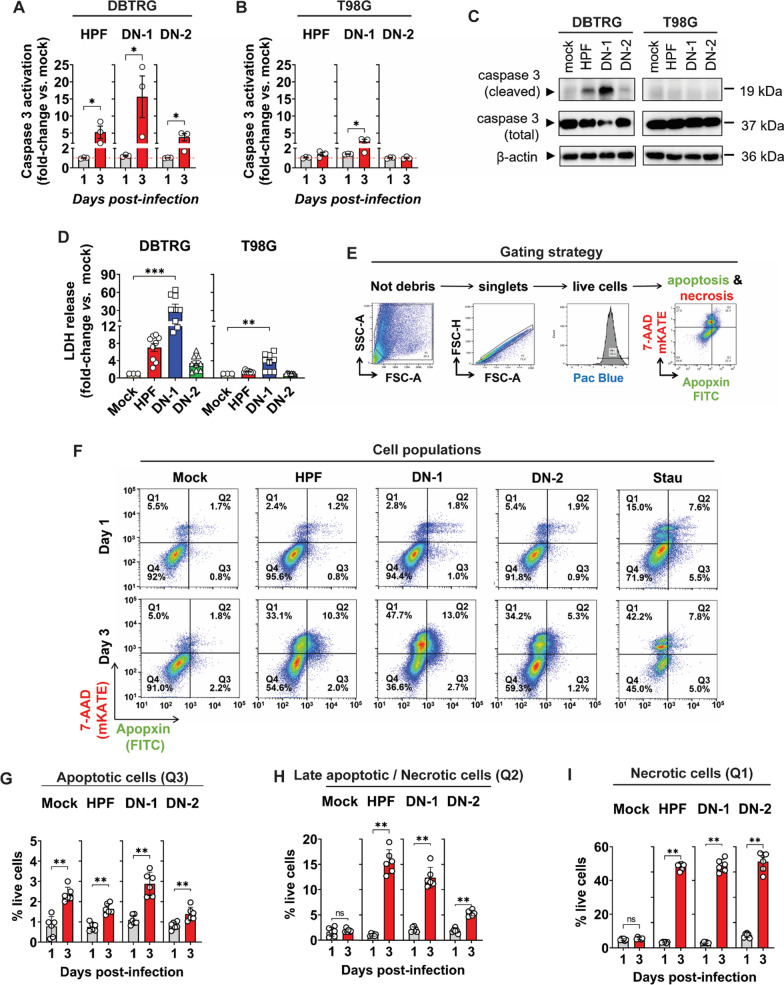
References
-
- Stupp R, Hegi ME, Mason WP, van den Bent MJ, Taphoorn MJ, Janzer RC, Ludwin SK, Allgeier A, Fisher B, Belanger K, Hau P, Brandes AA, Gijtenbeek J, Marosi C, Vecht CJ, Mokhtari K, Wesseling P, Villa S, Eisenhauer E, Gorlia T, Weller M, Lacombe D, Cairncross JG, Mirimanoff RO, Research European Organisation for, Tumour Treatment of Cancer Brain, Groups Radiation Oncology, and Group National Cancer Institute of Canada Clinical Trials. Effects of radiotherapy with concomitant and adjuvant temozolomide versus radiotherapy alone on survival in glioblastoma in a randomised phase iii study: 5-year analysis of the Eortc-Ncic Trial. Lancet Oncol. 2009;10(5):459–66. 10.1016/S1470-2045(09)70025-7. - DOI - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Research Materials
Miscellaneous
