

Biological basis of extensive pleiotropy between blood traits and cancer risk
- PMID: 38308367
- PMCID: PMC10837955
- DOI: 10.1186/s13073-024-01294-8
Biological basis of extensive pleiotropy between blood traits and cancer risk
Abstract
Background: The immune system has a central role in preventing carcinogenesis. Alteration of systemic immune cell levels may increase cancer risk. However, the extent to which common genetic variation influences blood traits and cancer risk remains largely undetermined. Here, we identify pleiotropic variants and predict their underlying molecular and cellular alterations.
Methods: Multivariate Cox regression was used to evaluate associations between blood traits and cancer diagnosis in cases in the UK Biobank. Shared genetic variants were identified from the summary statistics of the genome-wide association studies of 27 blood traits and 27 cancer types and subtypes, applying the conditional/conjunctional false-discovery rate approach. Analysis of genomic positions, expression quantitative trait loci, enhancers, regulatory marks, functionally defined gene sets, and bulk- and single-cell expression profiles predicted the biological impact of pleiotropic variants. Plasma small RNAs were sequenced to assess association with cancer diagnosis.
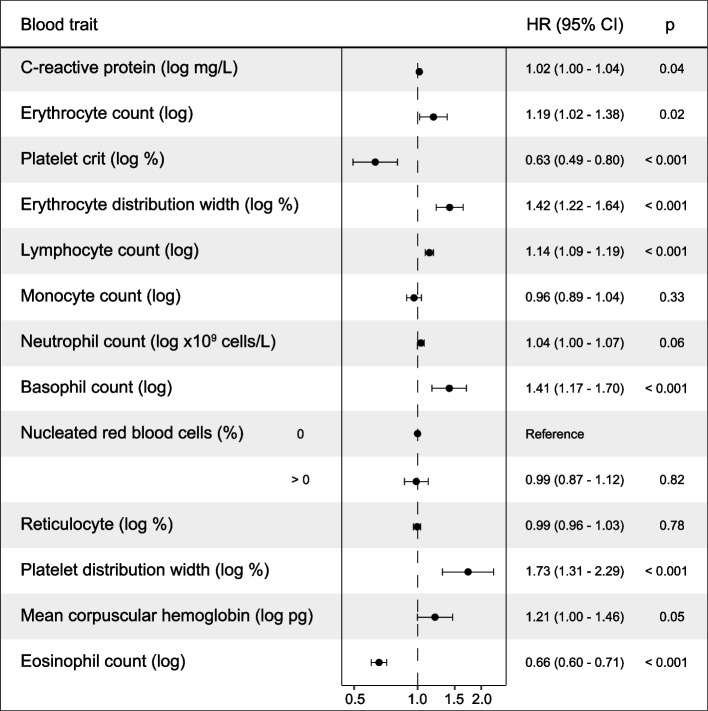
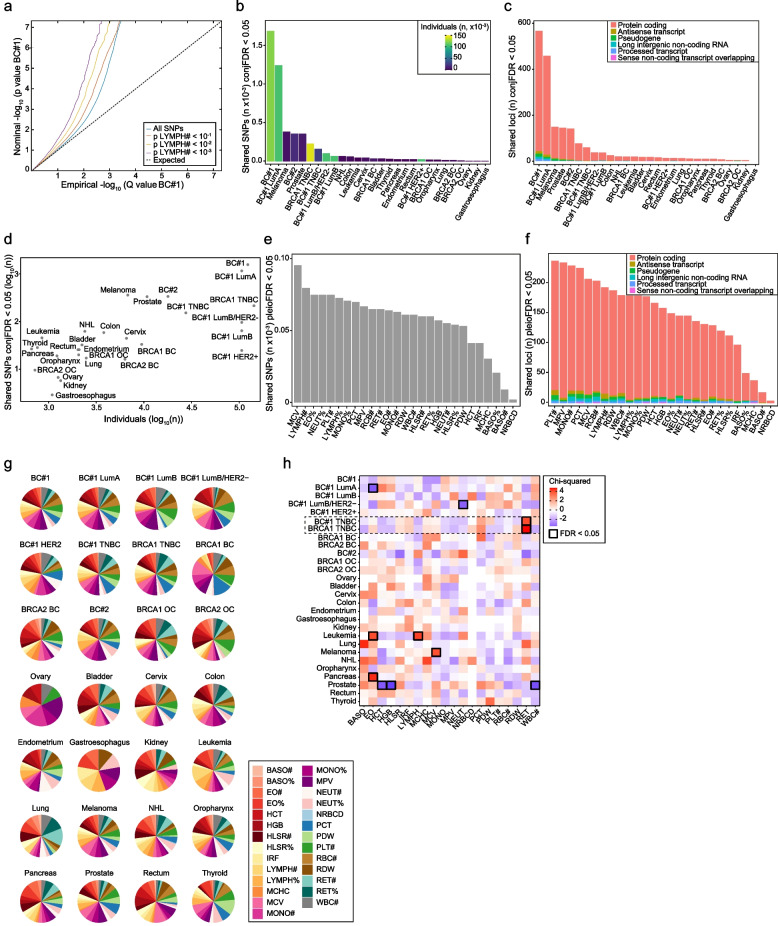
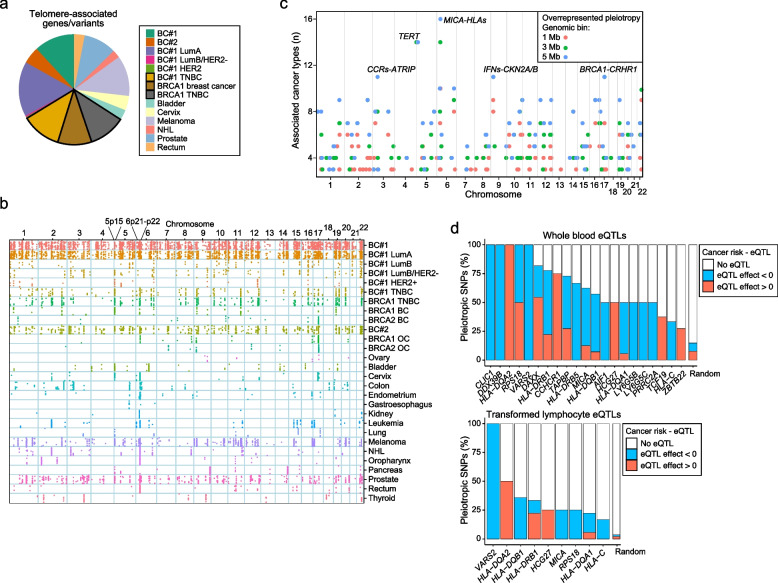
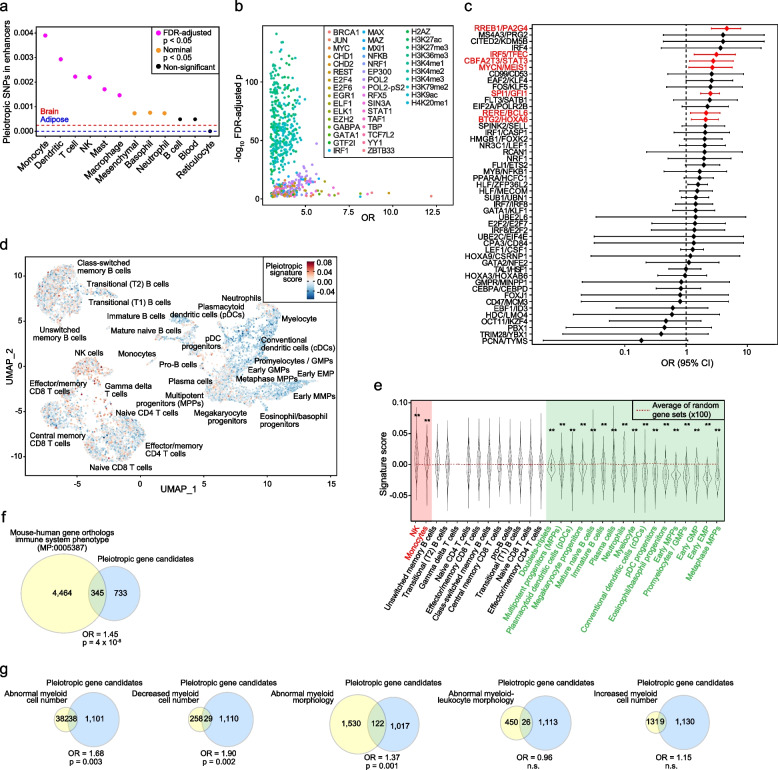
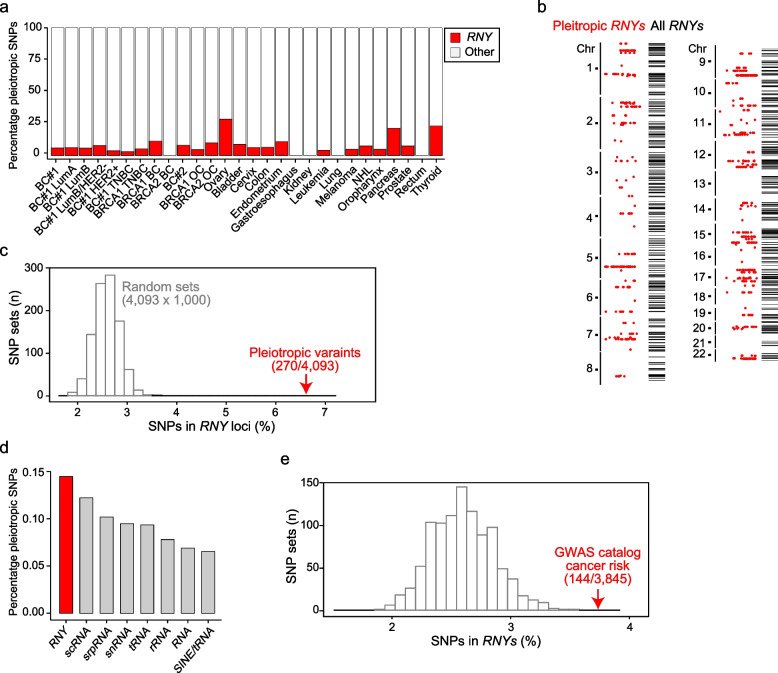
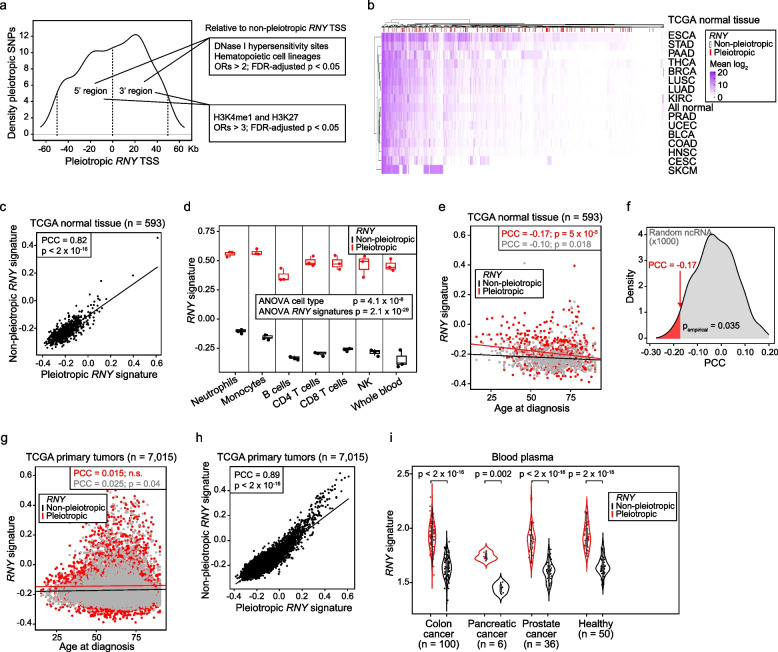
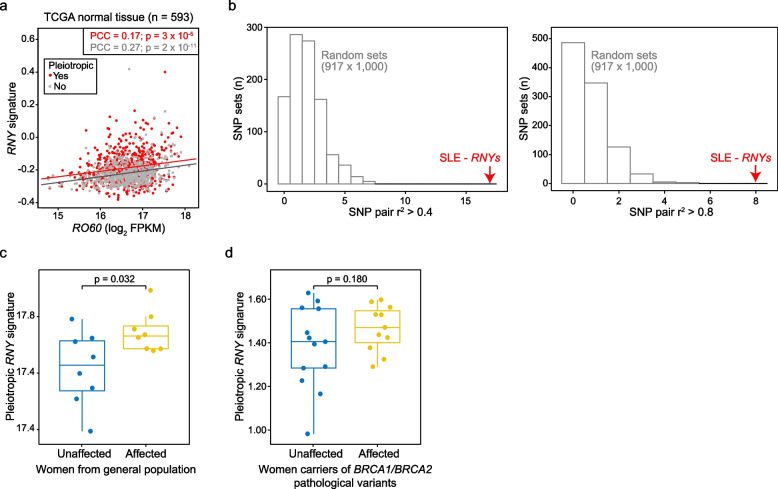
Results: The study identified 4093 common genetic variants, involving 1248 gene loci, that contributed to blood-cancer pleiotropism. Genomic hotspots of pleiotropism include chromosomal regions 5p15-TERT and 6p21-HLA. Genes whose products are involved in regulating telomere length are found to be enriched in pleiotropic variants. Pleiotropic gene candidates are frequently linked to transcriptional programs that regulate hematopoiesis and define progenitor cell states of immune system development. Perturbation of the myeloid lineage is indicated by pleiotropic associations with defined master regulators and cell alterations. Eosinophil count is inversely associated with cancer risk. A high frequency of pleiotropic associations is also centered on the regulation of small noncoding Y-RNAs. Predicted pleiotropic Y-RNAs show specific regulatory marks and are overabundant in the normal tissue and blood of cancer patients. Analysis of plasma small RNAs in women who developed breast cancer indicates there is an overabundance of Y-RNA preceding neoplasm diagnosis.
Conclusions: This study reveals extensive pleiotropism between blood traits and cancer risk. Pleiotropism is linked to factors and processes involved in hematopoietic development and immune system function, including components of the major histocompatibility complexes, and regulators of telomere length and myeloid lineage. Deregulation of Y-RNAs is also associated with pleiotropism. Overexpression of these elements might indicate increased cancer risk.
Keywords: Blood trait; Cancer; Eosinophil; Hematopoiesis; Myeloid; Pleiotropy; Telomere; Y-RNA.
© 2024. The Author(s).
Conflict of interest statement
The authors declare that they have no competing interests.
Figures
References
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Research Materials
