Oxidative stress changes interactions between 2 bacterial species from competitive to facilitative
- PMID: 38315734
- PMCID: PMC10881020
- DOI: 10.1371/journal.pbio.3002482
Oxidative stress changes interactions between 2 bacterial species from competitive to facilitative
Abstract
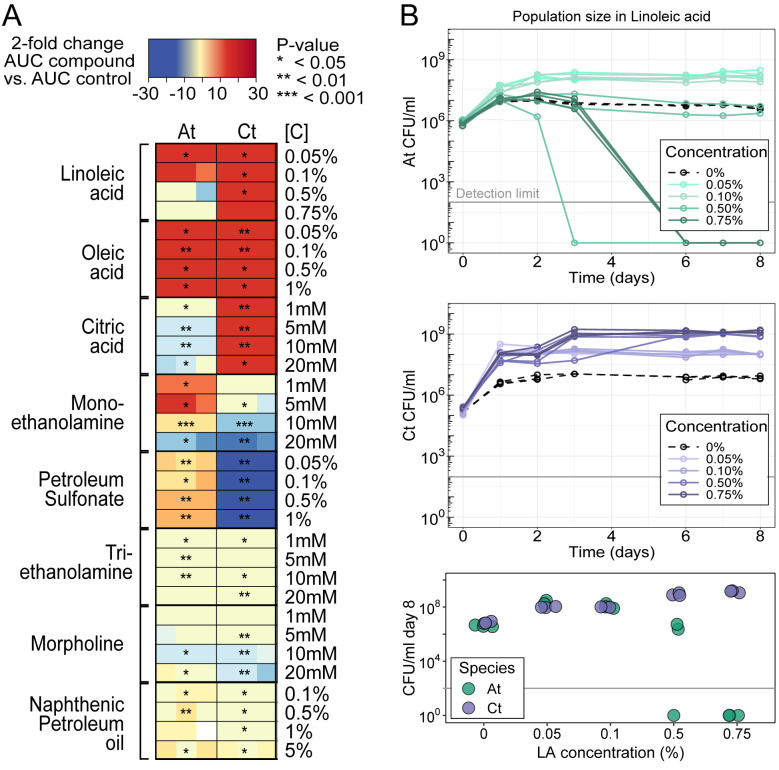
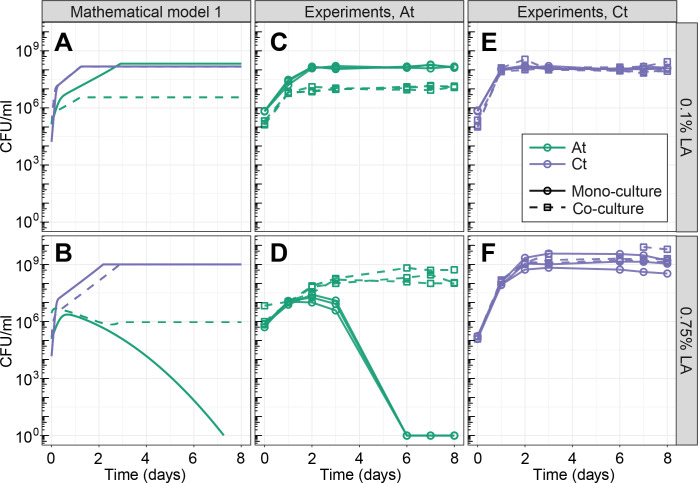
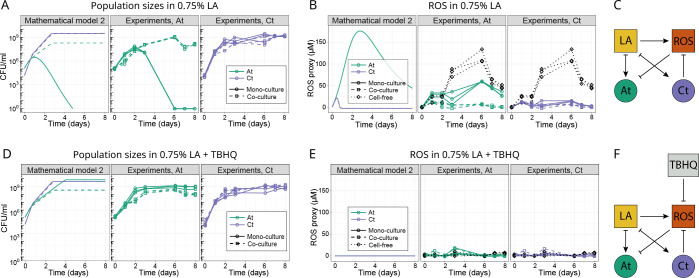
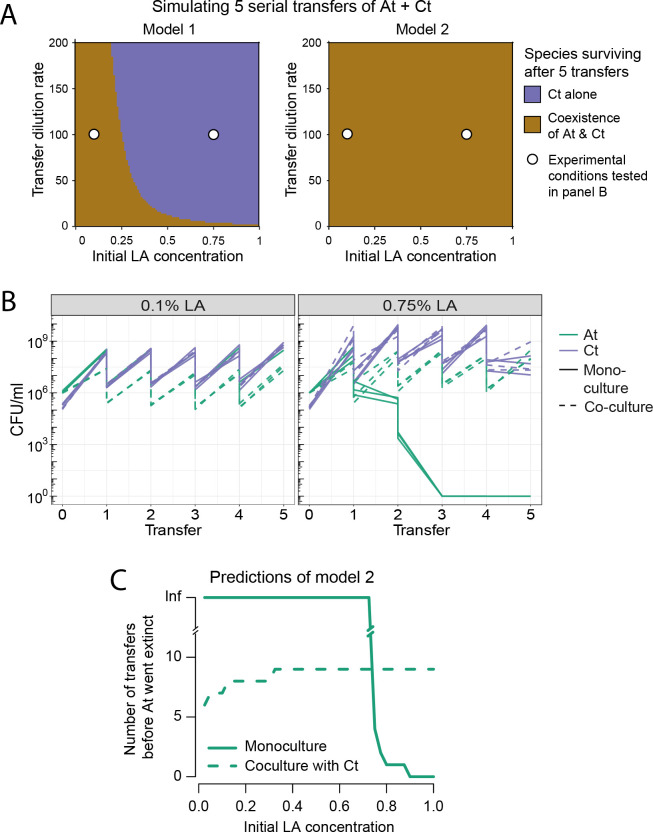
Knowing how species interact within microbial communities is crucial to predicting and controlling community dynamics, but interactions can depend on environmental conditions. The stress-gradient hypothesis (SGH) predicts that species are more likely to facilitate each other in harsher environments. Even if the SGH gives some intuition, quantitative modeling of the context-dependency of interactions requires understanding the mechanisms behind the SGH. In this study, we show with both experiments and a theoretical analysis that varying the concentration of a single compound, linoleic acid (LA), modifies the interaction between 2 bacterial species, Agrobacterium tumefaciens and Comamonas testosteroni, from competitive at a low concentration, to facilitative at higher concentrations where LA becomes toxic for one of the 2 species. We demonstrate that the mechanism behind facilitation is that one species is able to reduce reactive oxygen species (ROS) that are produced spontaneously at higher concentrations of LA, allowing for short-term rescue of the species that is sensitive to ROS and longer coexistence in serial transfers. In our system, competition and facilitation between species can occur simultaneously, and changing the concentration of a single compound can alter the balance between the two.
Copyright: © 2024 Di Martino et al. This is an open access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures