

Efficient prime editing in two-cell mouse embryos using PEmbryo
- PMID: 38321114
- PMCID: PMC11631759
- DOI: 10.1038/s41587-023-02106-x
Efficient prime editing in two-cell mouse embryos using PEmbryo
Abstract
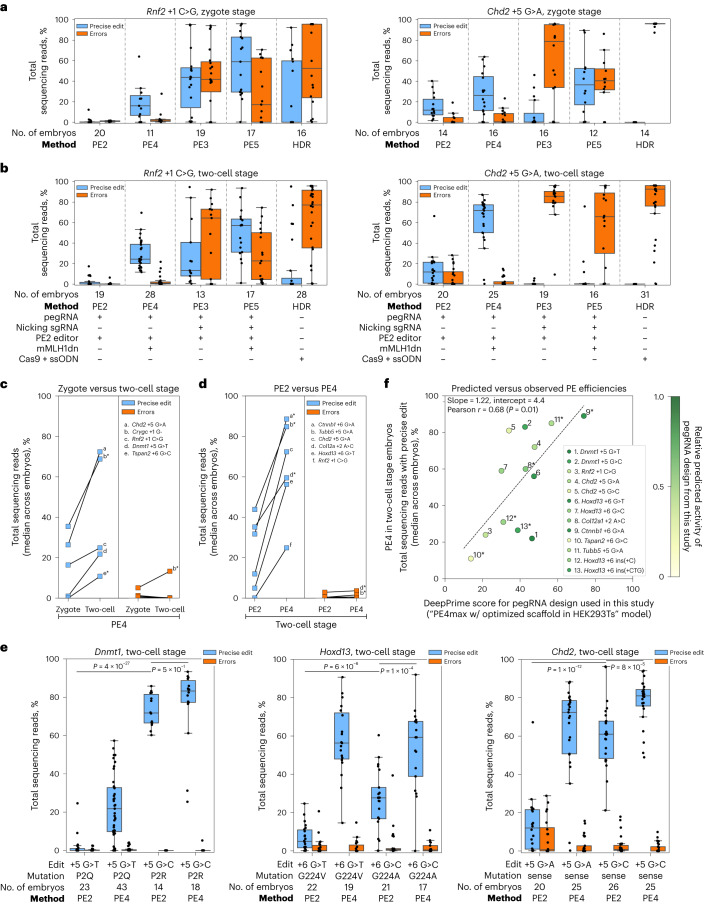
Using transient inhibition of DNA mismatch repair during a permissive stage of development, we demonstrate highly efficient prime editing of mouse embryos with few unwanted, local byproducts (average 58% precise edit frequency, 0.5% on-target error frequency across 13 substitution edits at 8 sites), enabling same-generation phenotyping of founders. Whole-genome sequencing reveals that mismatch repair inhibition increases off-target indels at low-complexity regions in the genome without any obvious phenotype in mice.
© 2024. The Author(s).
Conflict of interest statement
Competing interests: B.A. is an advisory board member with options for Arbor Biotechnologies and Tessera Therapeutics. B.A. holds equity in Celsius Therapeutics. D.R.L. is a consultant and equity holder of Beam Therapeutics, Prime Medicine, Pairwise Plants, Chroma Medicine, Resonance Medicine, Exo Therapeutics and Nvelop Therapeutics, companies that use genome editing or engineering. P.J.C. is currently an employee of Prime Medicine. J.E.T. is an advisor to Nereid Therapeutics and Prolific Machines. E.A.I. has pending interest in VUS Diagnostics. B.A., D.R.L. and P.J.C. are co-inventors on prime editing patent applications filed through their respective institutions. The other authors declare no competing interests.
Figures


References
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
