

The vaccinia chondroitin sulfate binding protein drives host membrane curvature to facilitate fusion
- PMID: 38321165
- PMCID: PMC10933376
- DOI: 10.1038/s44319-023-00040-2
The vaccinia chondroitin sulfate binding protein drives host membrane curvature to facilitate fusion
Abstract
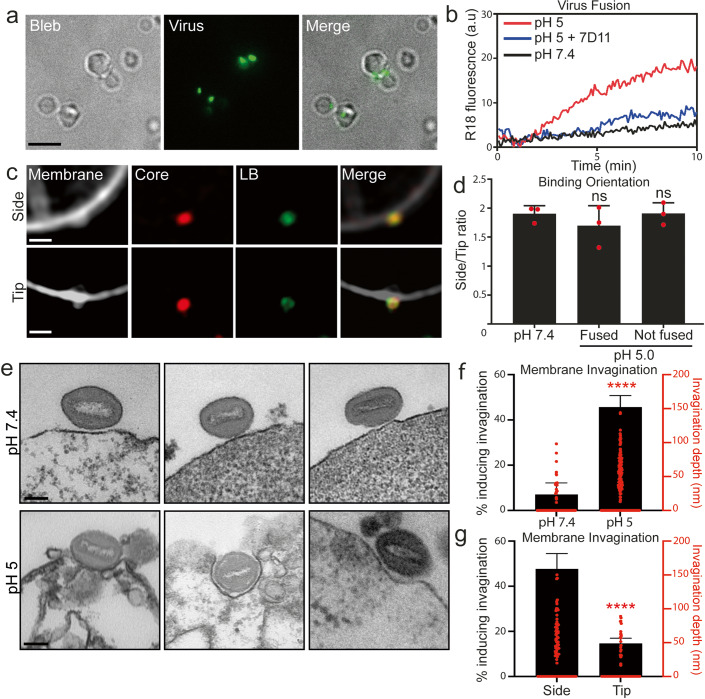
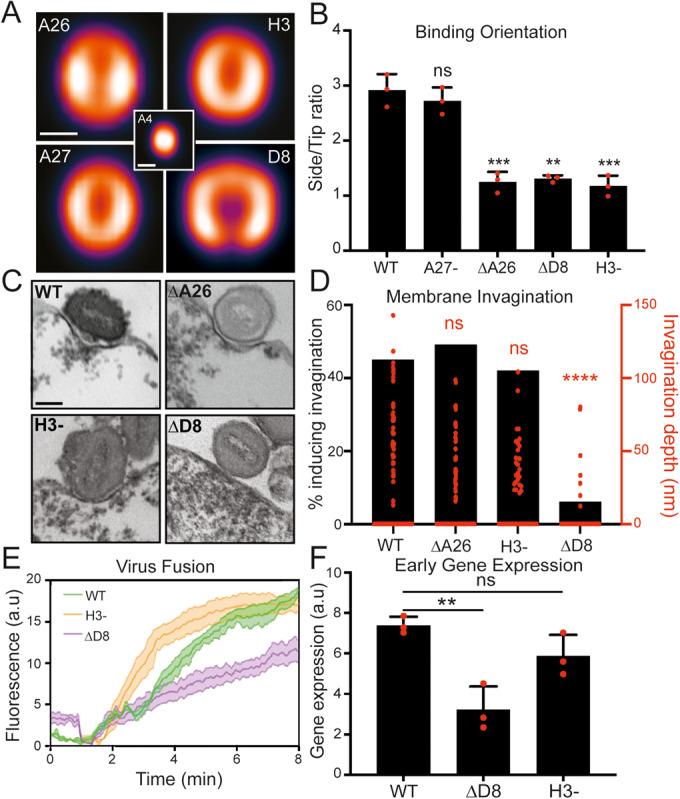
Cellular attachment of viruses determines their cell tropism and species specificity. For entry, vaccinia, the prototypic poxvirus, relies on four binding proteins and an eleven-protein entry fusion complex. The contribution of the individual virus binding proteins to virion binding orientation and membrane fusion is unclear. Here, we show that virus binding proteins guide side-on virion binding and promote curvature of the host membrane towards the virus fusion machinery to facilitate fusion. Using a membrane-bleb model system together with super-resolution and electron microscopy we find that side-bound vaccinia virions induce membrane invagination in the presence of low pH. Repression or deletion of individual binding proteins reveals that three of four contribute to binding orientation, amongst which the chondroitin sulfate binding protein, D8, is required for host membrane bending. Consistent with low-pH dependent macropinocytic entry of vaccinia, loss of D8 prevents virion-associated macropinosome membrane bending, disrupts fusion pore formation and infection. Our results show that viral binding proteins are active participants in successful virus membrane fusion and illustrate the importance of virus protein architecture for successful infection.
Keywords: Glycosaminoglycans; Membrane Bending; Membrane Fusion; Poxvirus; Virus Entry.
© 2024. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures





References
-
- Aldaz-carroll L, Whitbeck JC, De Leon MP, Lou H, Hirao L, Isaacs SN, Moss B, Eisenberg RJ, Cohen GH. Epitope-mapping studies define two major neutralization sites on the vaccinia virus extracellular enveloped virus. J Virol. 2005;79:6260–6271. doi: 10.1128/JVI.79.10.6260-6271.2005. - DOI - PMC - PubMed
-
- Banfield BW, Leduc Y, Esford L, Schubert K, Tufaro F. Sequential isolation of proteoglycan synthesis mutants by using herpes simplex virus as a selective agent: evidence for a proteoglycan-independent virus entry pathway. J Virol. 1995;69:3290–3298. doi: 10.1128/jvi.69.6.3290-3298.1995. - DOI - PMC - PubMed
-
- Bergefall K, Trybala E, Johansson M, Uyama T, Naito S, Yamada S, Kitagawa H, Sugahara K, Bergstro T. Chondroitin sulfate characterized by the E-disaccharide unit is a potent inhibitor of herpes simplex virus infectivity and provides the virus binding sites on gro2C cells. J Biol Chem. 2005;280:32193–32199. doi: 10.1074/jbc.M503645200. - DOI - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
