

Epithelial zonation along the mouse and human small intestine defines five discrete metabolic domains
- PMID: 38321203
- PMCID: PMC11654995
- DOI: 10.1038/s41556-023-01337-z
Epithelial zonation along the mouse and human small intestine defines five discrete metabolic domains
Abstract
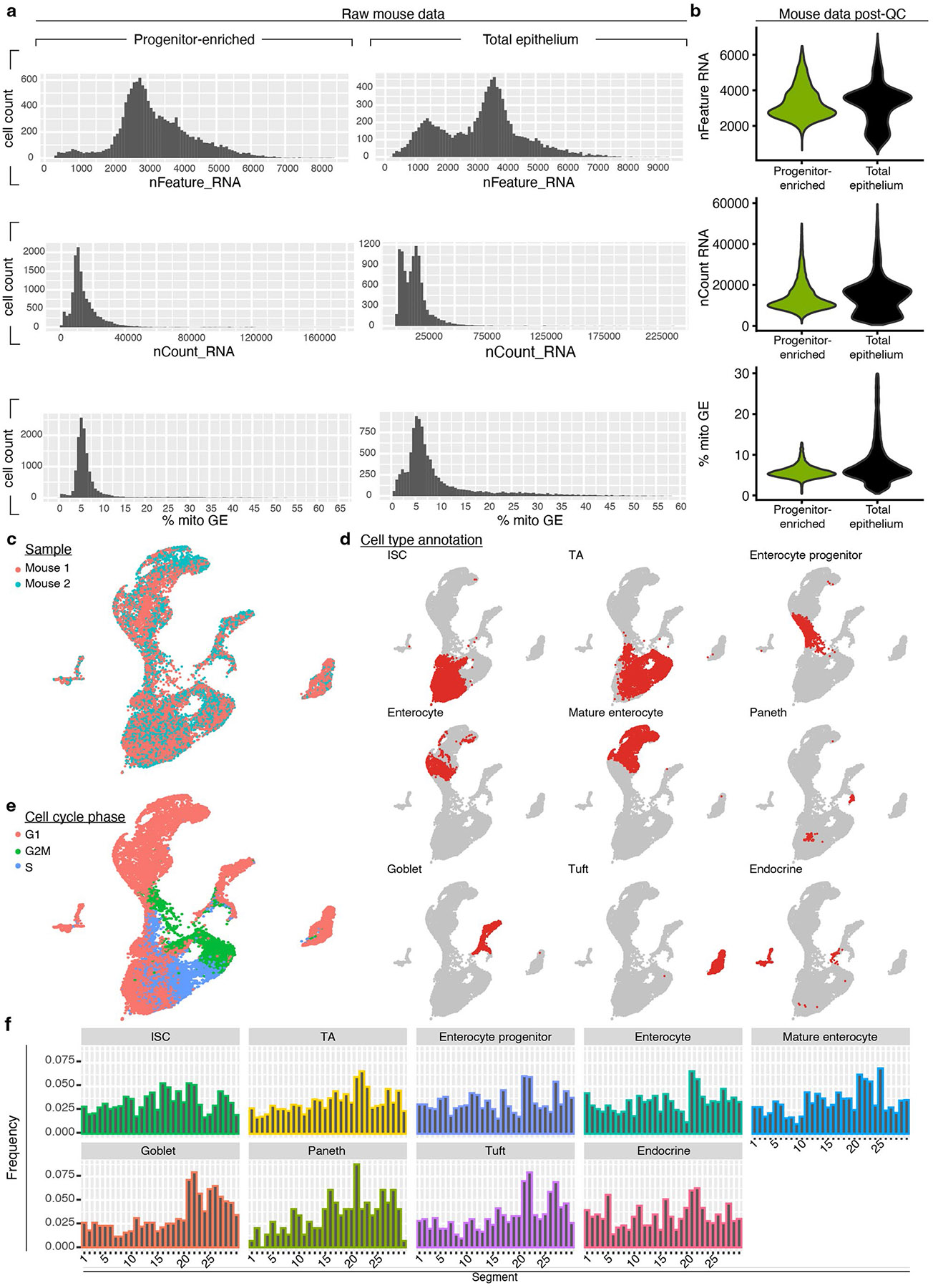
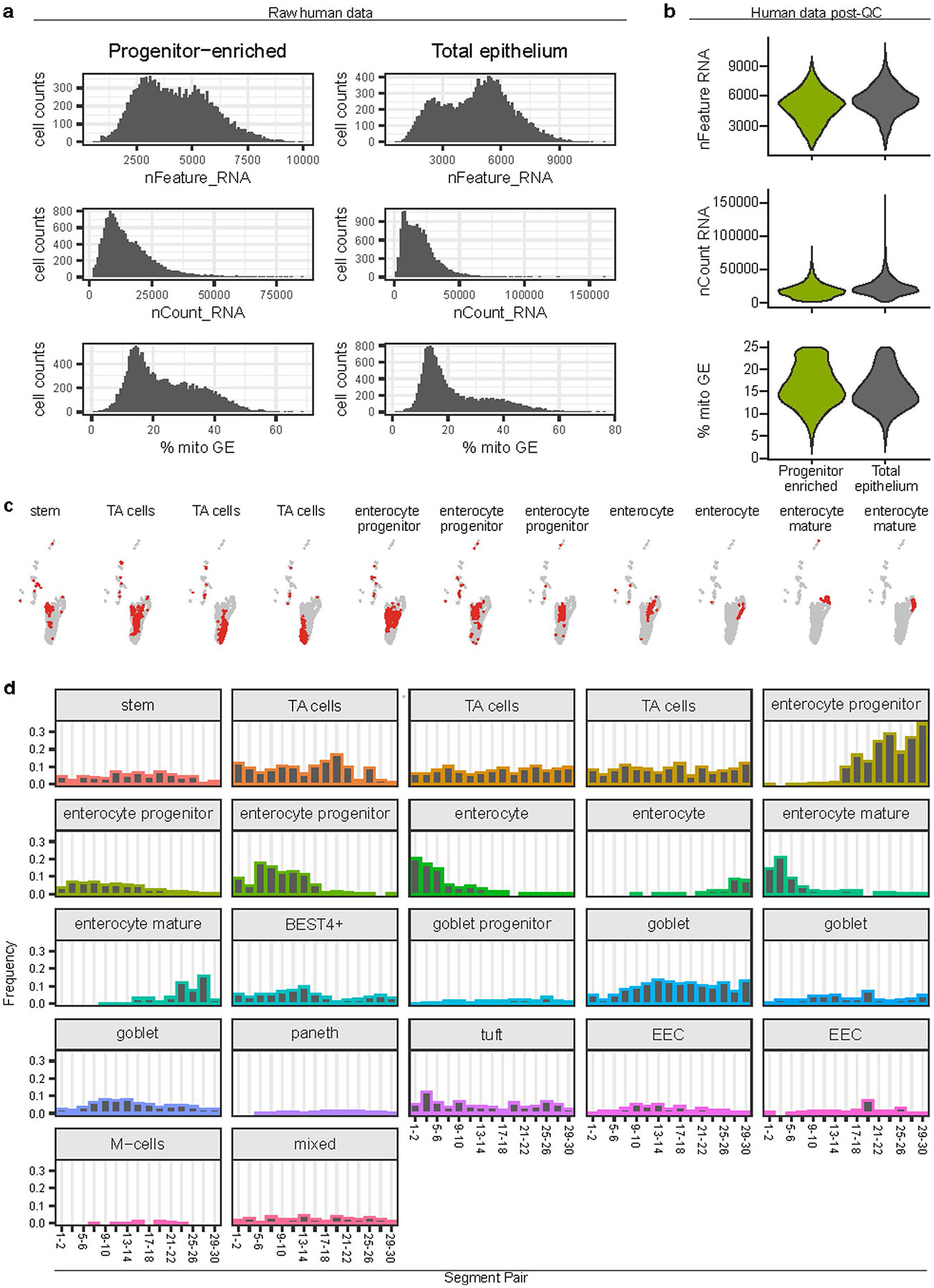
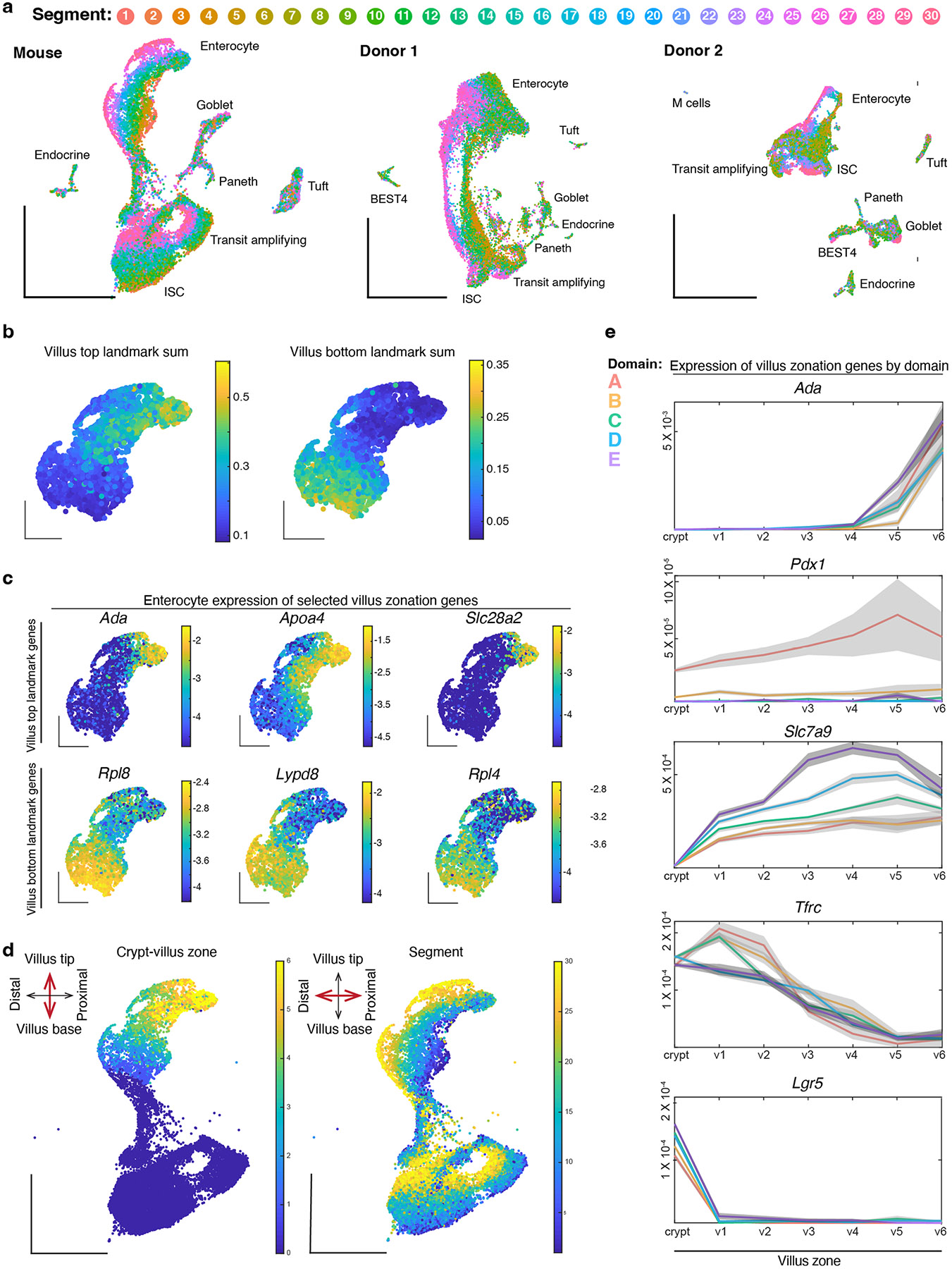
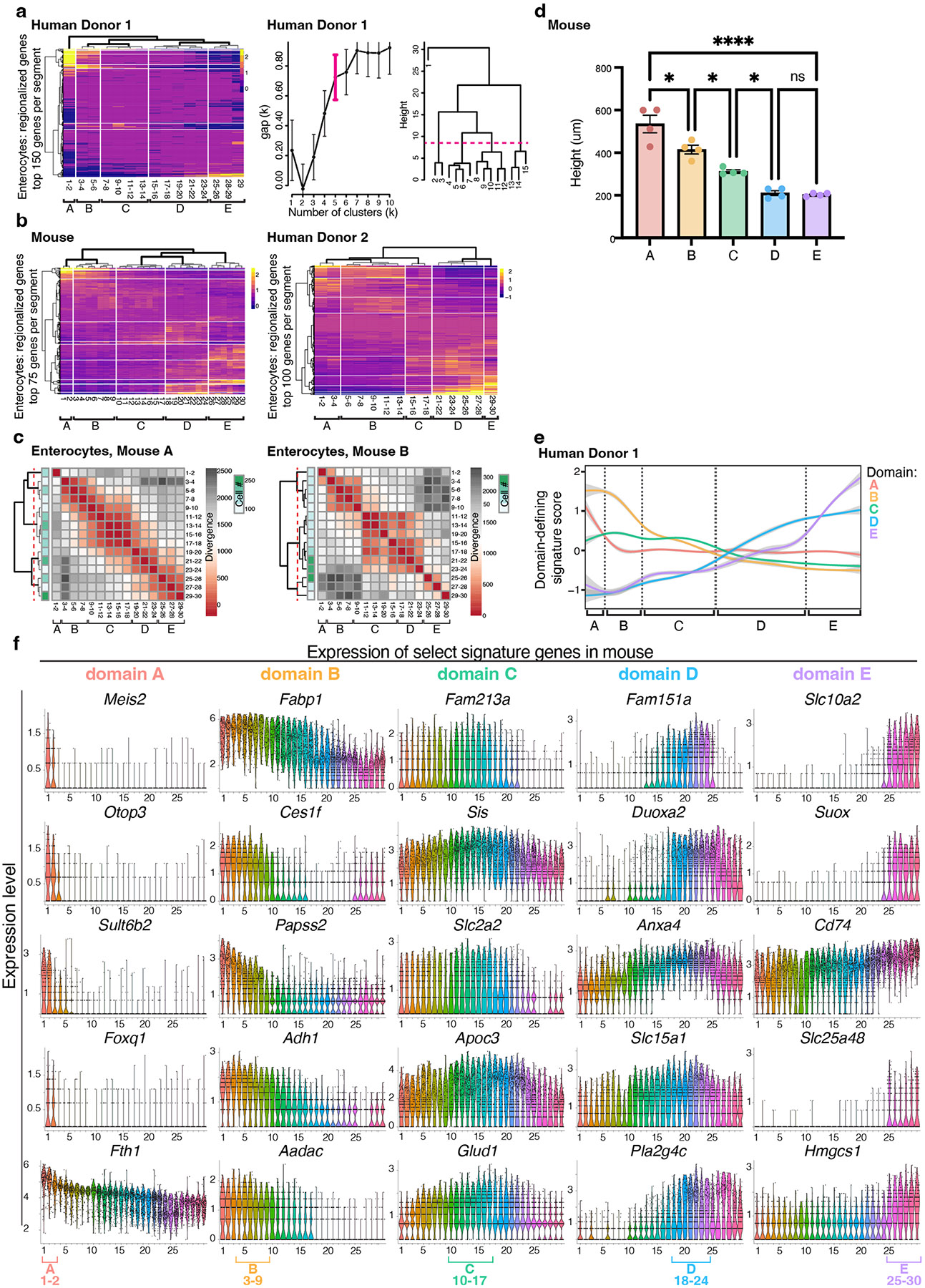
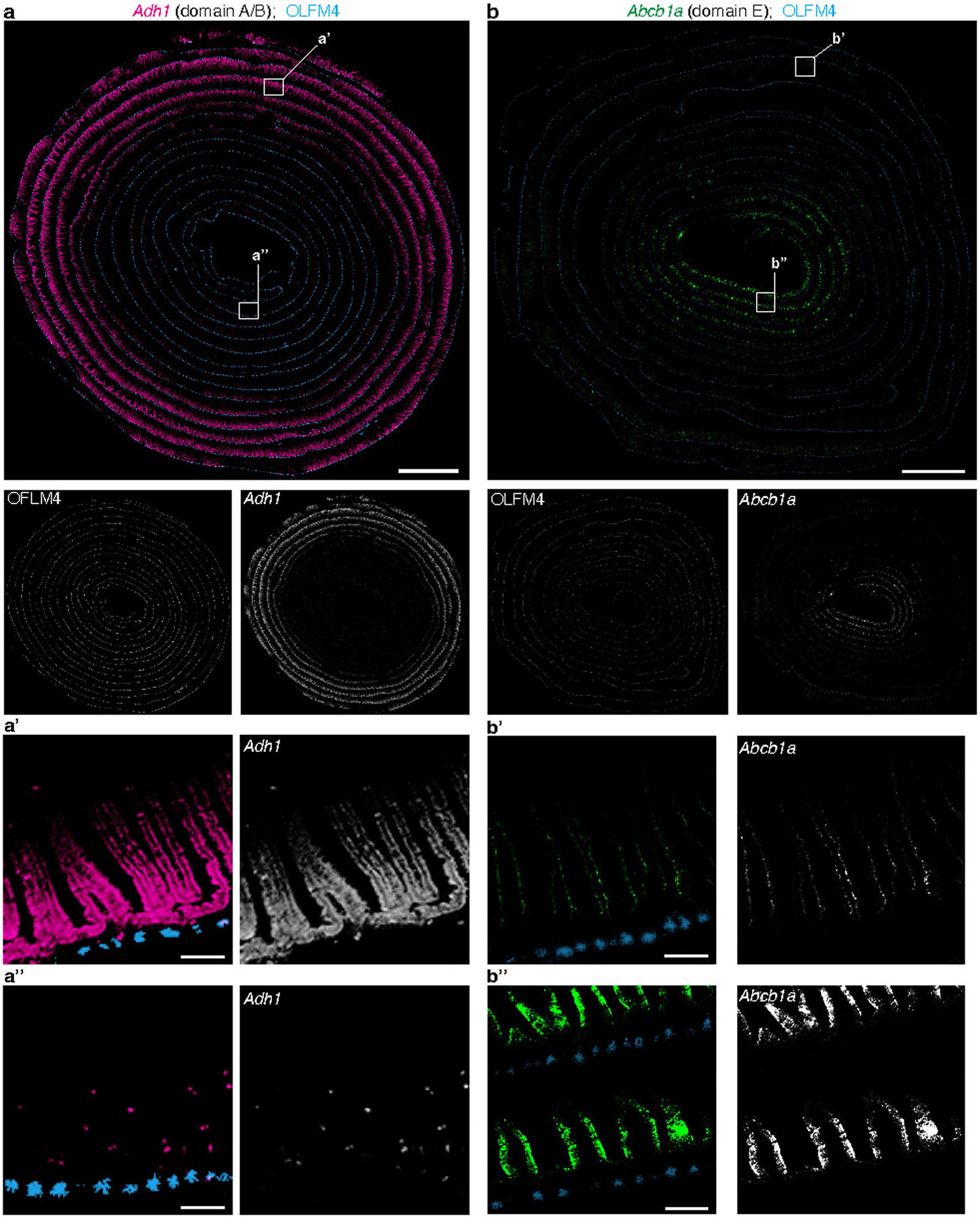
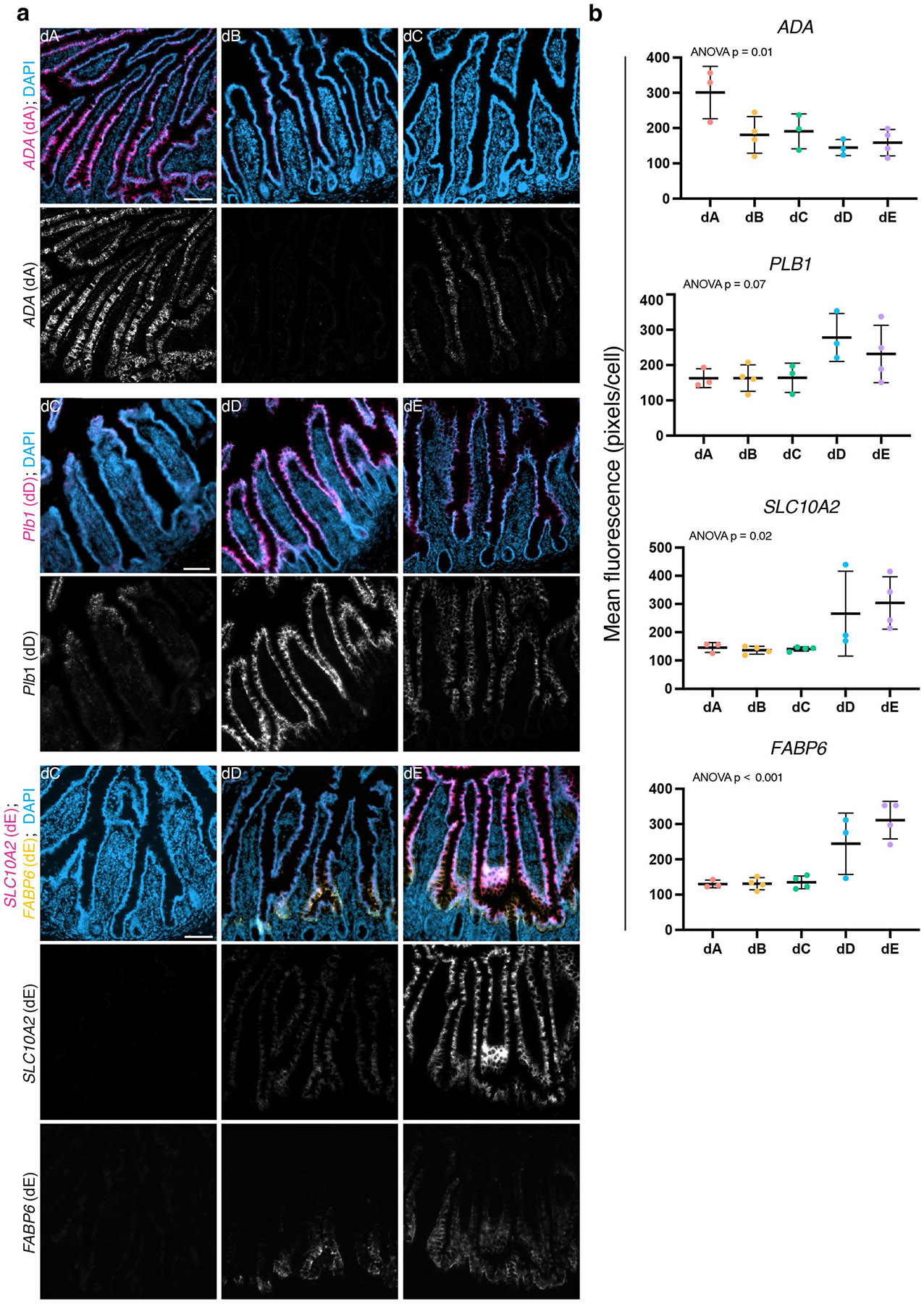
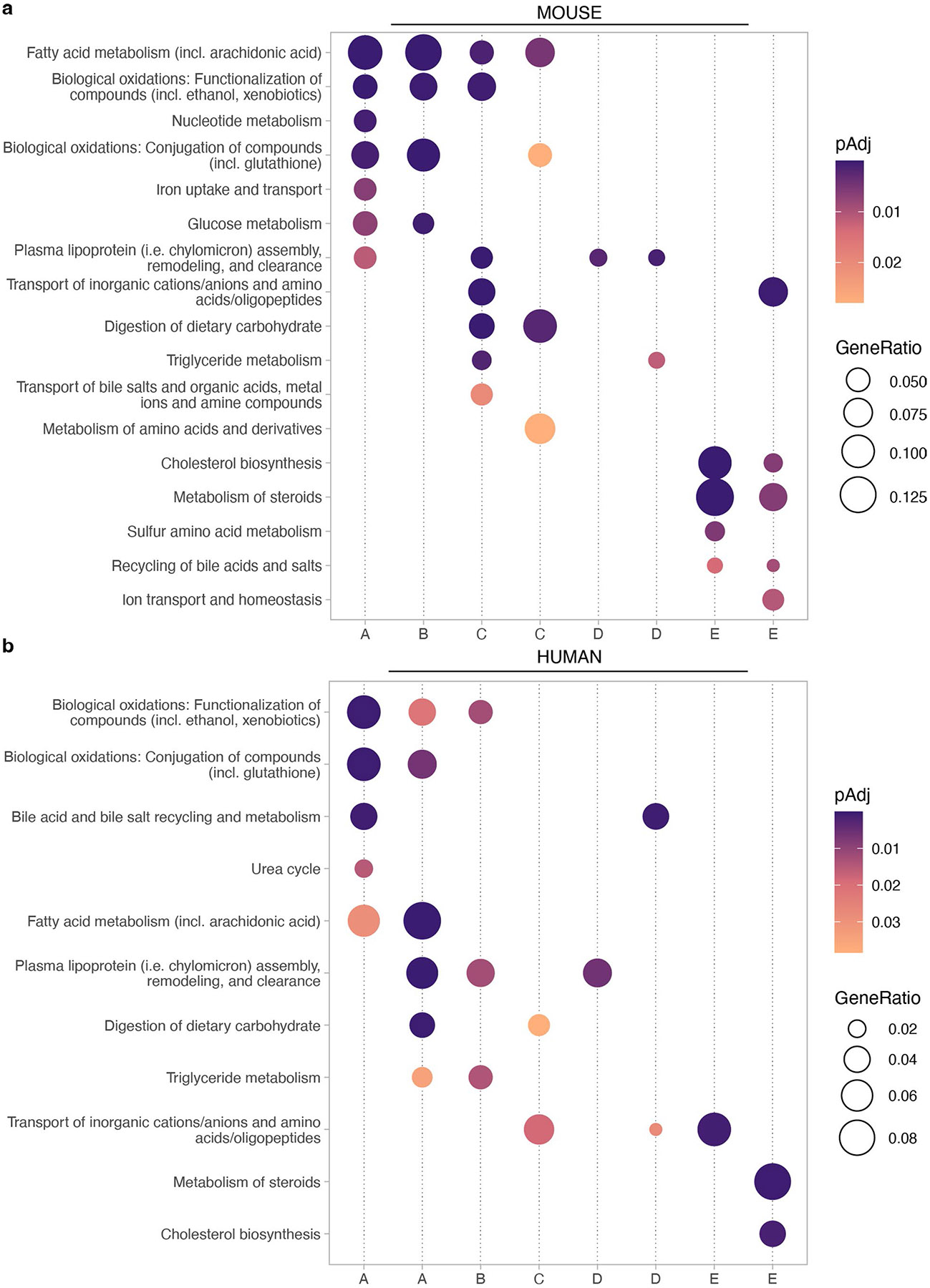
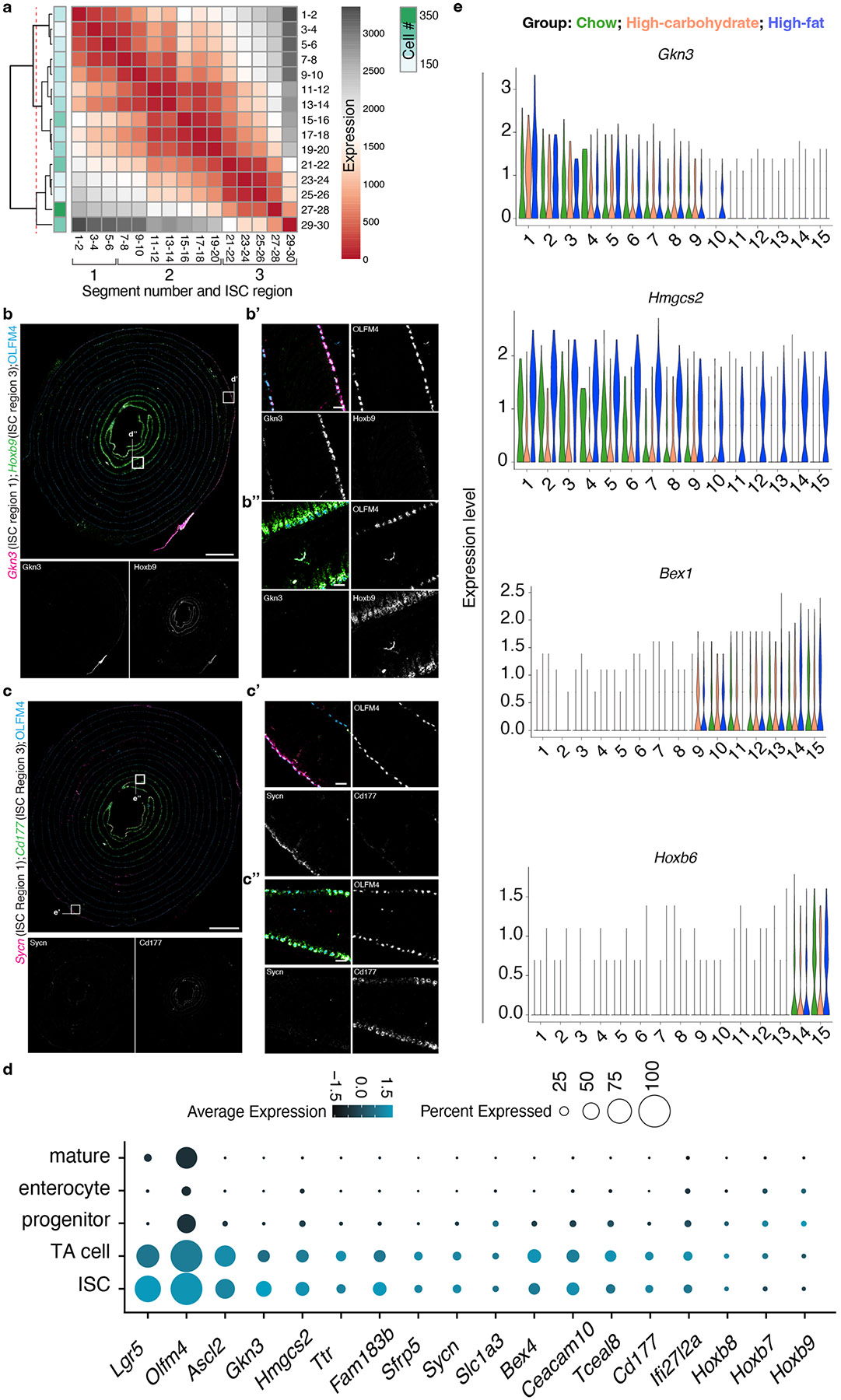
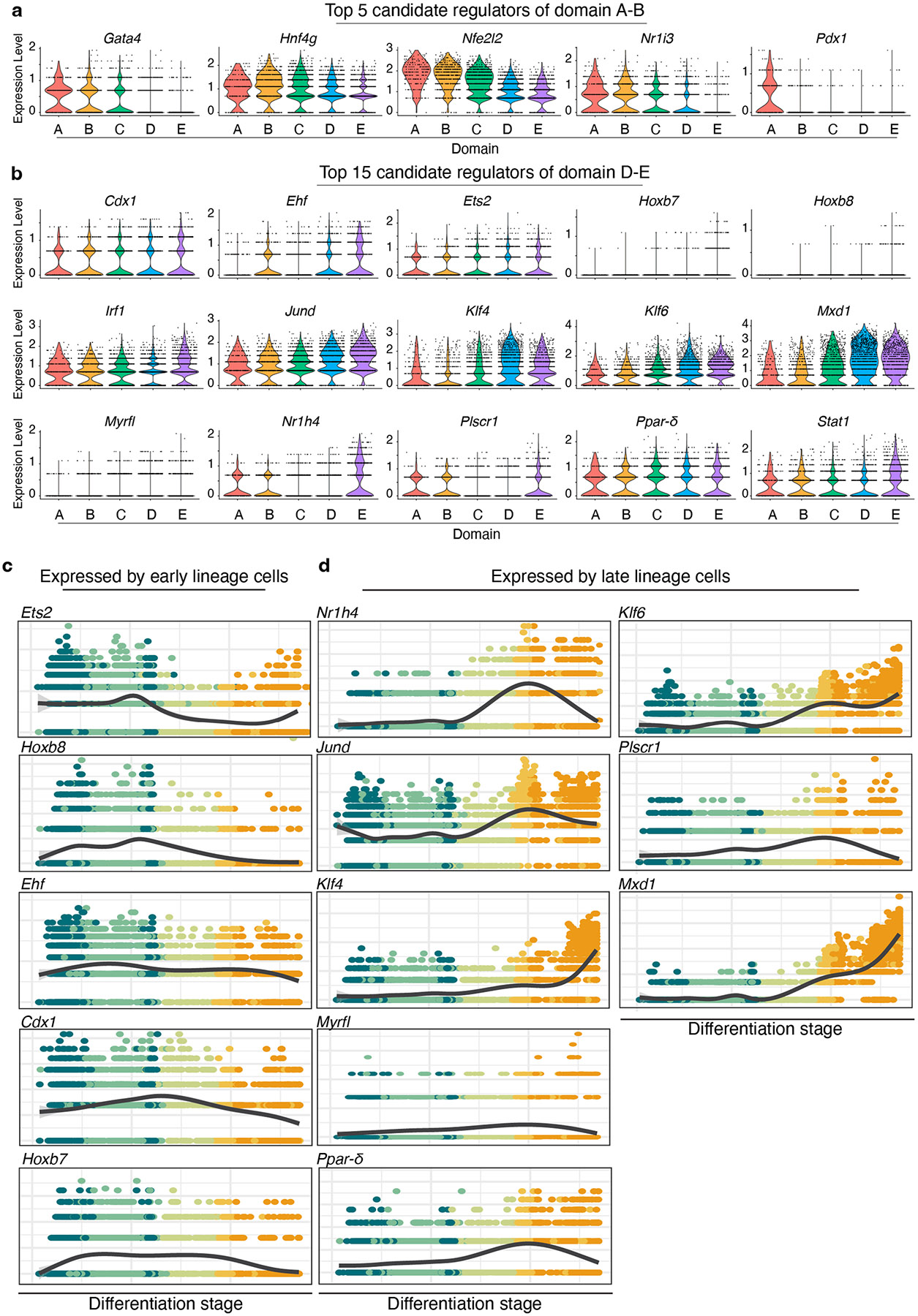
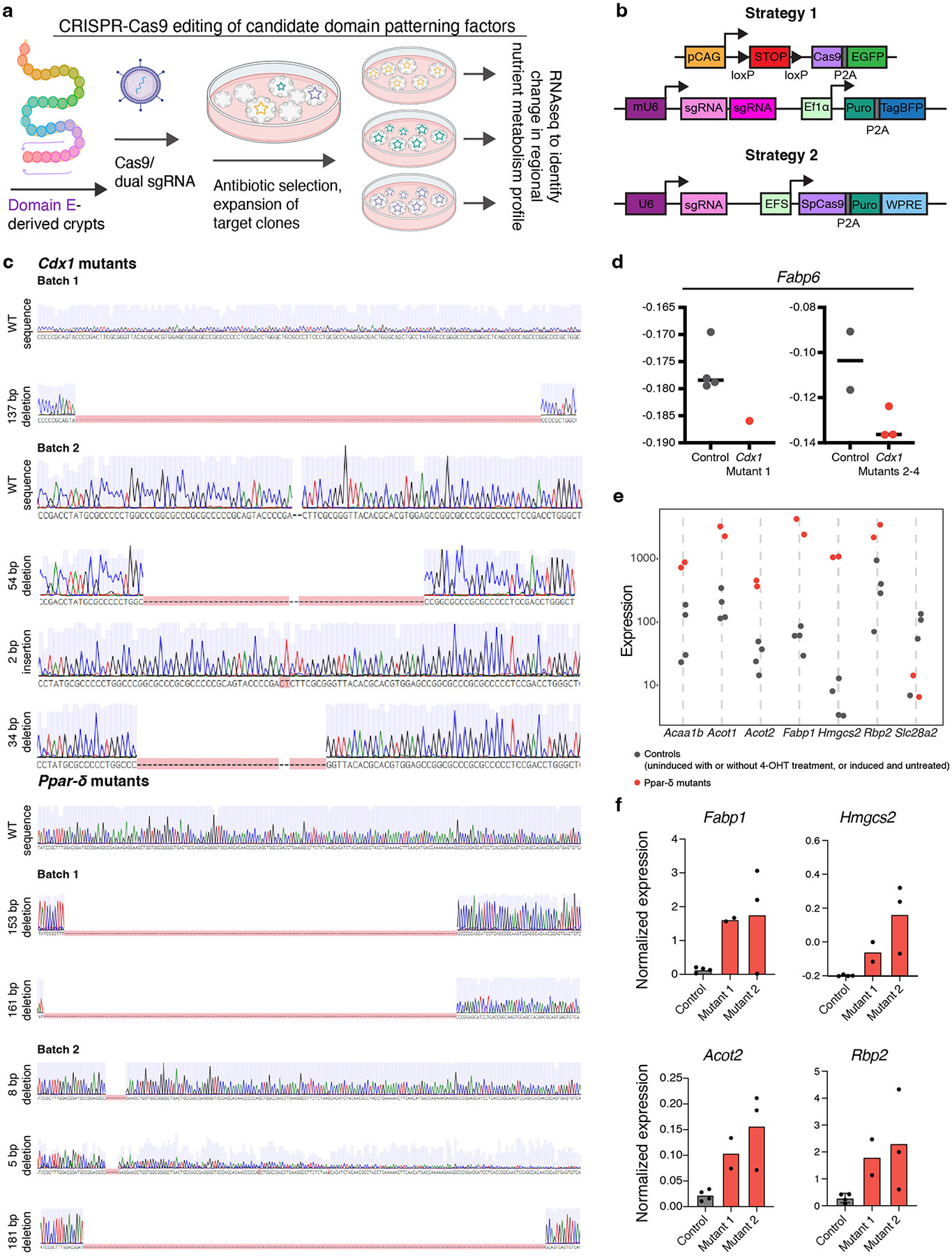
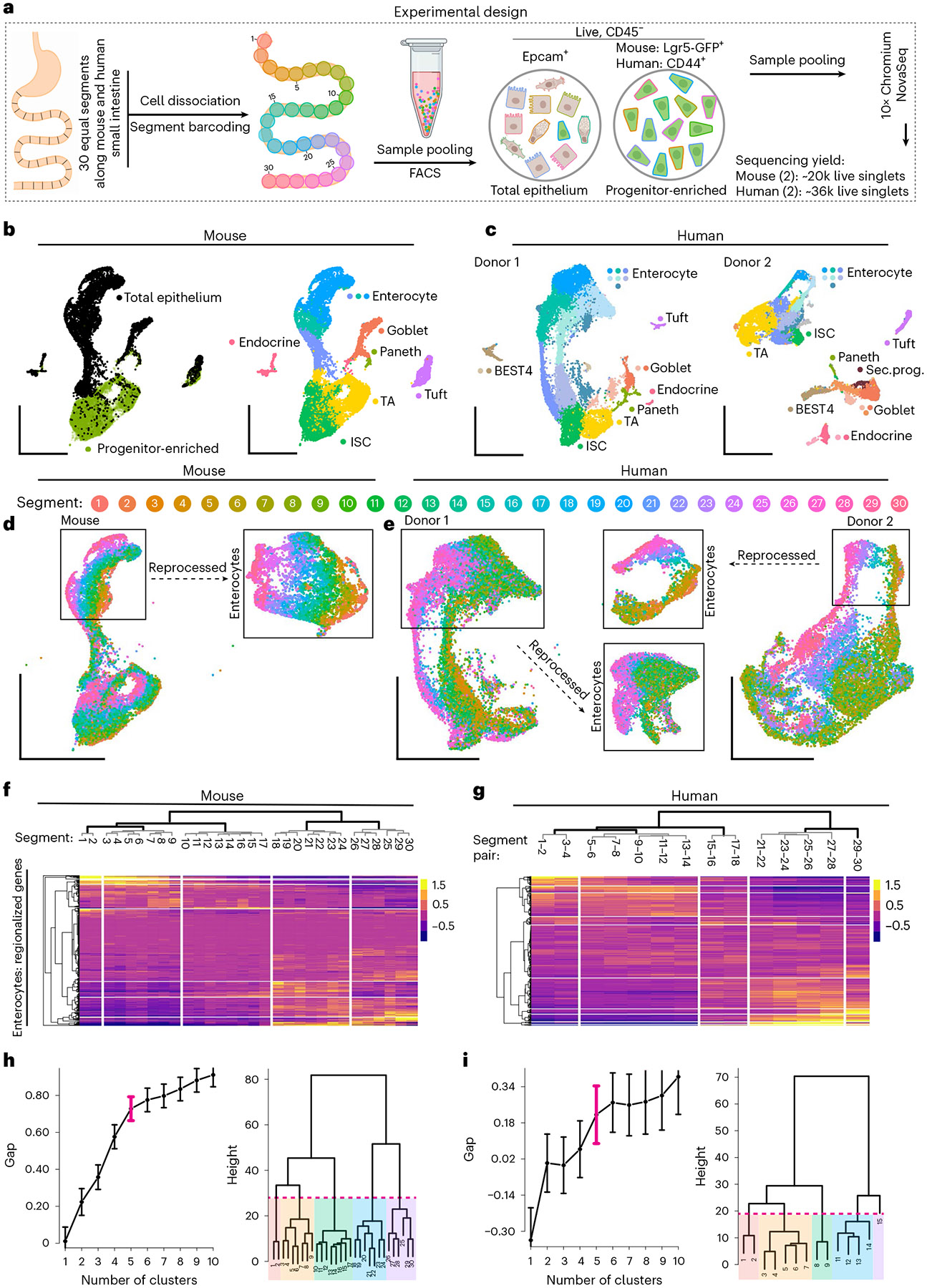
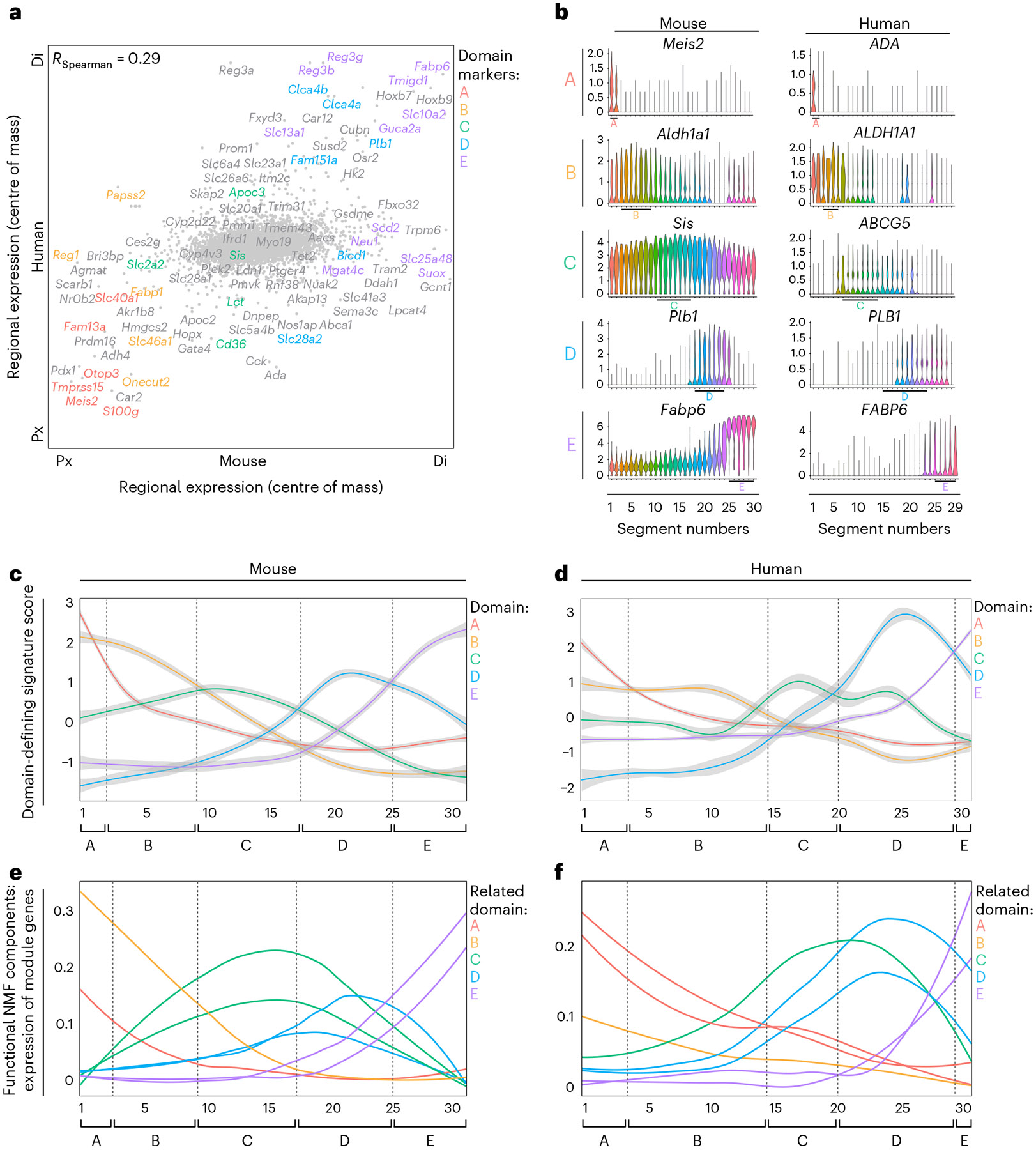
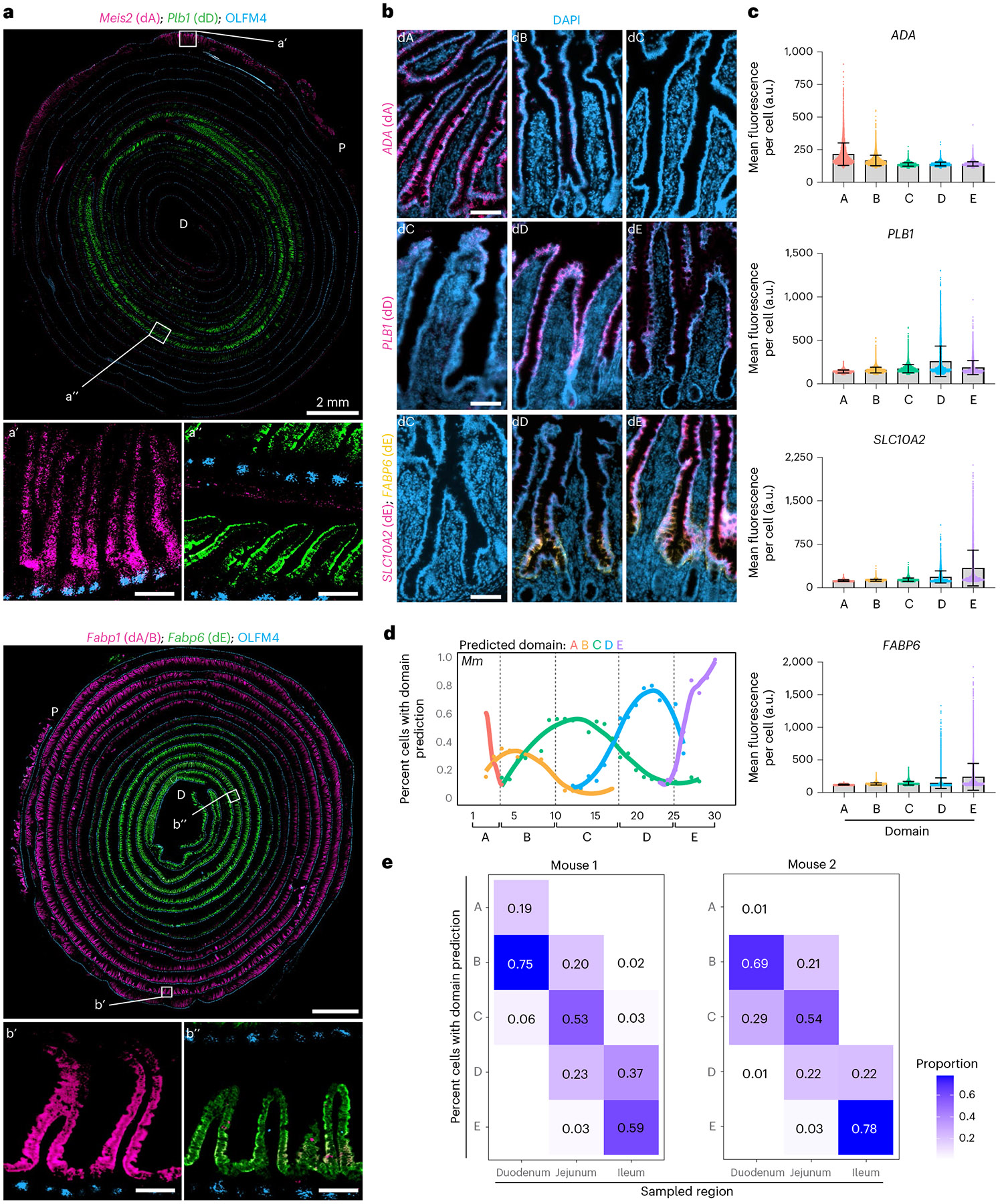
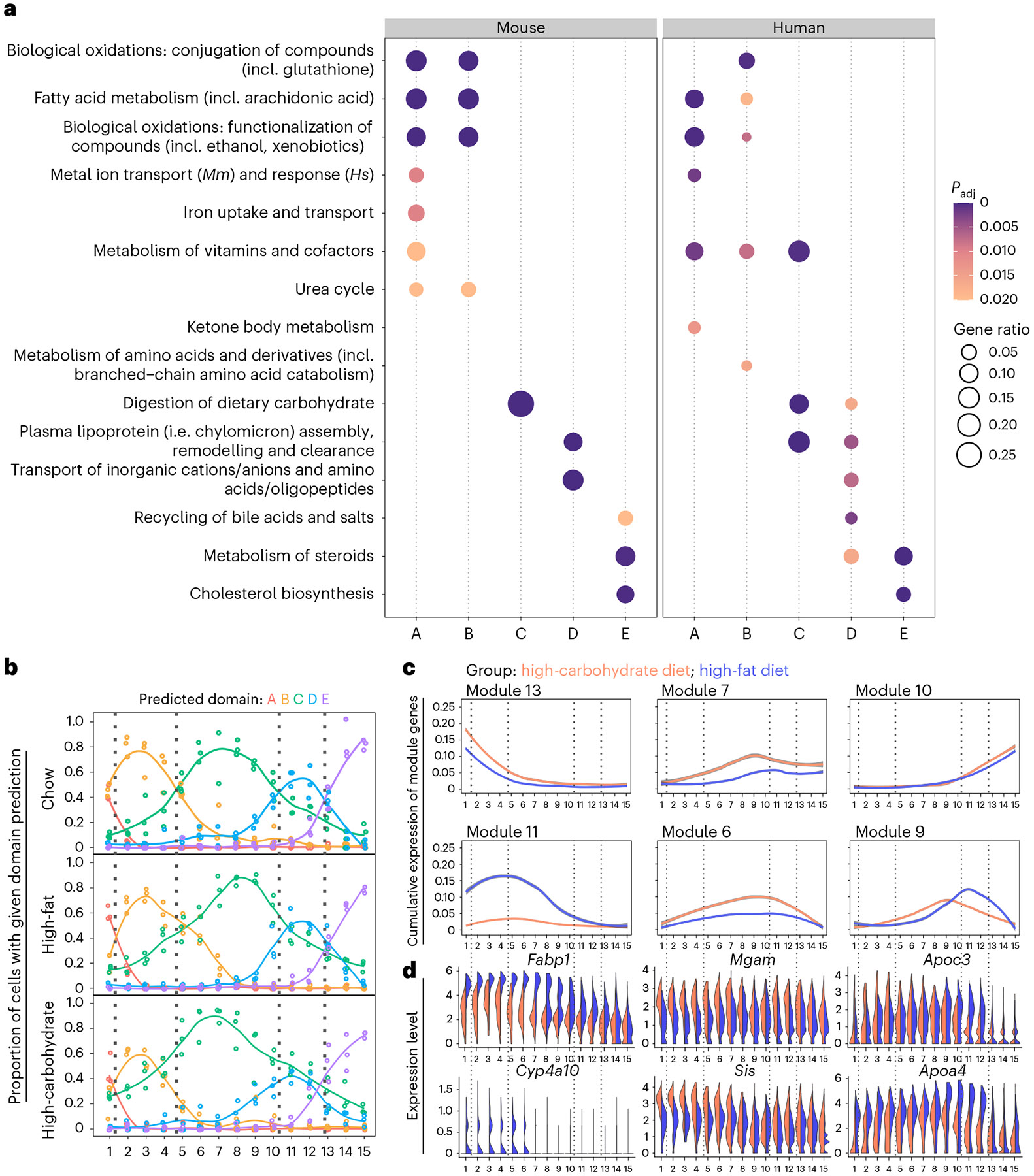
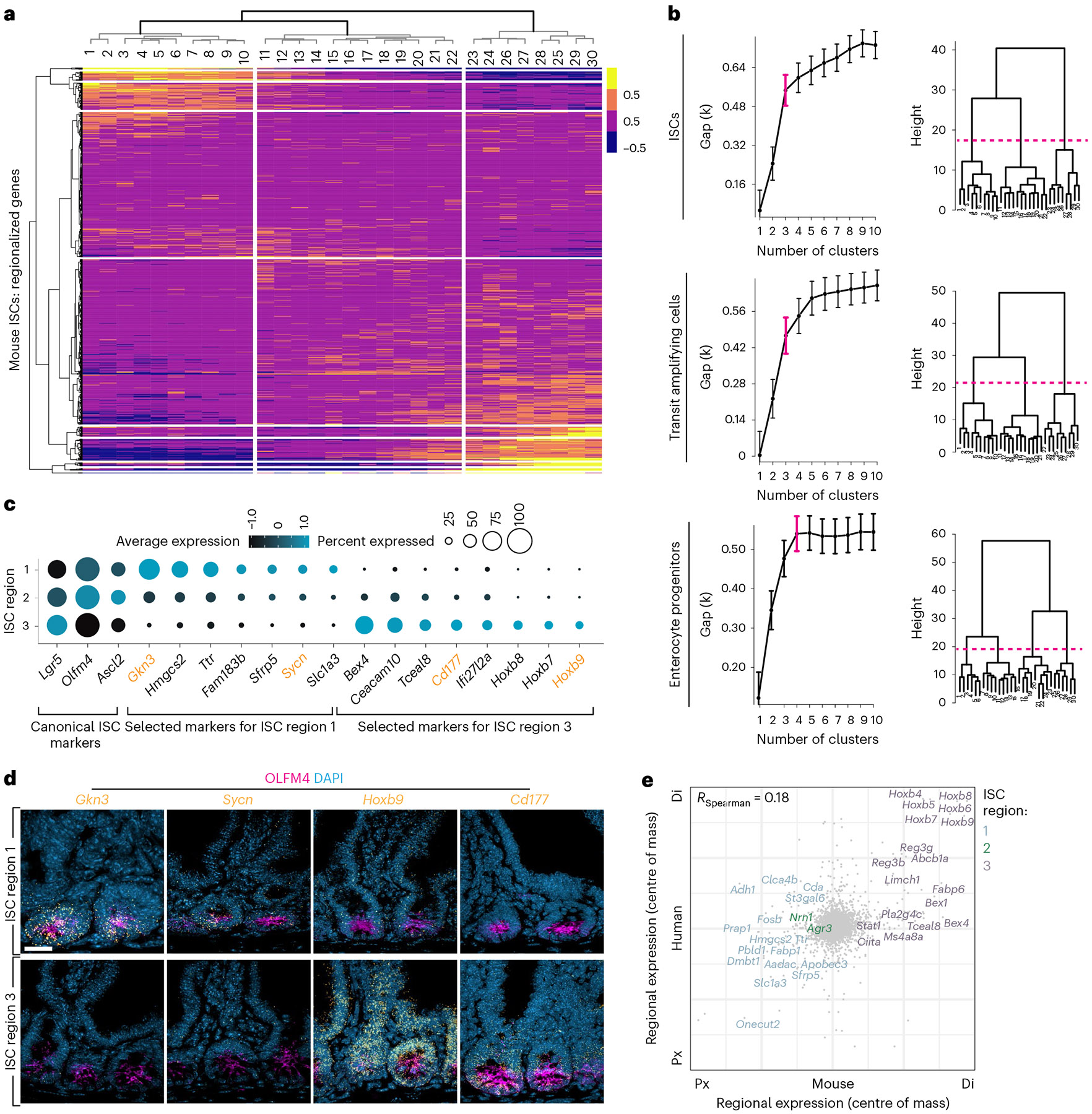
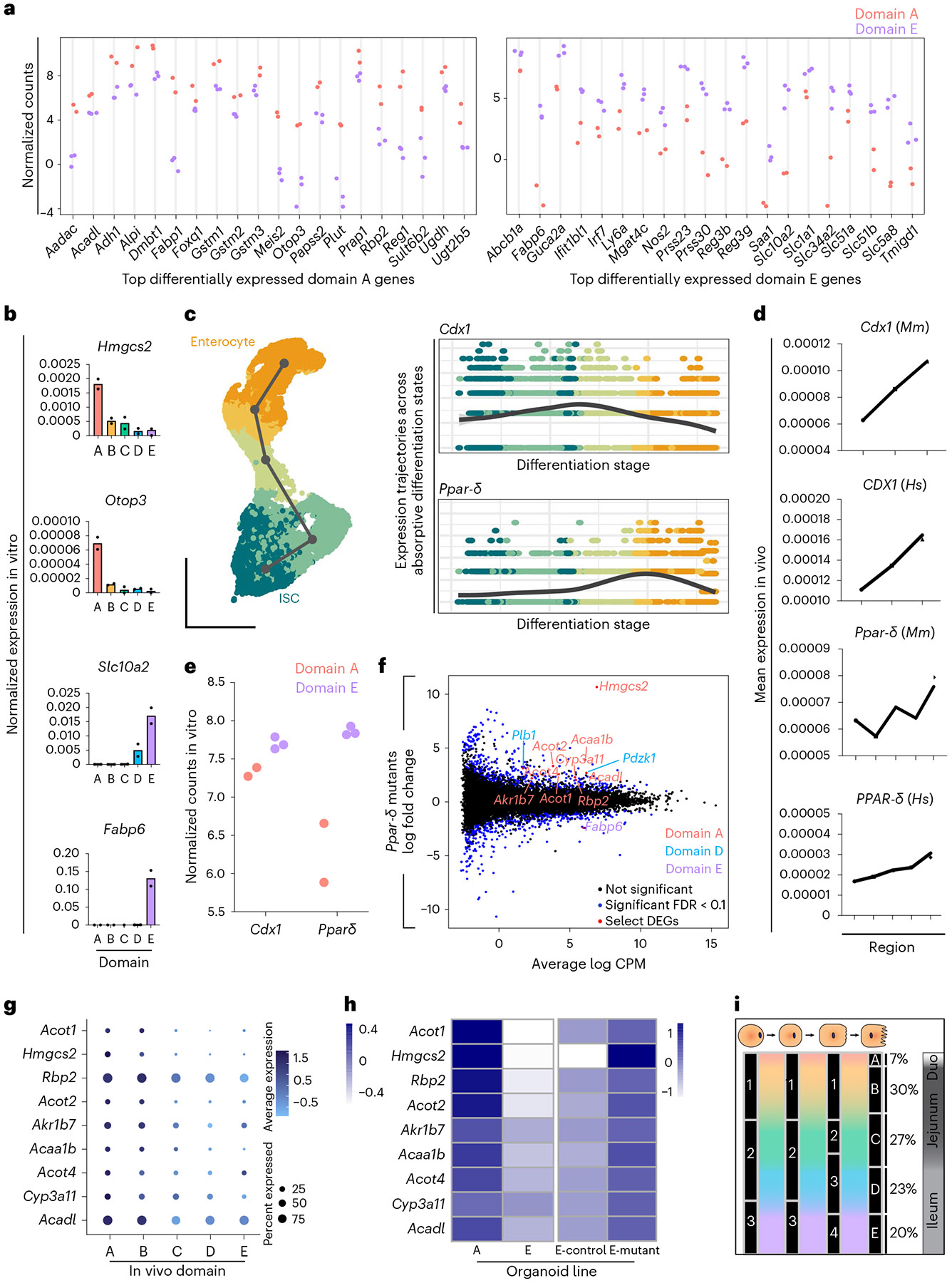
A key aspect of nutrient absorption is the exquisite division of labour across the length of the small intestine, with individual nutrients taken up at different proximal:distal positions. For millennia, the small intestine was thought to comprise three segments with indefinite borders: the duodenum, jejunum and ileum. By examining the fine-scale longitudinal transcriptional patterns that span the mouse and human small intestine, we instead identified five domains of nutrient absorption that mount distinct responses to dietary changes, and three regional stem cell populations. Molecular domain identity can be detected with machine learning, which provides a systematic method to computationally identify intestinal domains in mice. We generated a predictive model of transcriptional control of domain identity and validated the roles of Ppar-δ and Cdx1 in patterning lipid metabolism-associated genes. These findings represent a foundational framework for the zonation of absorption across the mammalian small intestine.
© 2024. The Author(s), under exclusive licence to Springer Nature Limited.
Figures
Update of
-
Epithelial zonation along the mouse and human small intestine defines five discrete metabolic domains.bioRxiv [Preprint]. 2023 Sep 22:2023.09.20.558726. doi: 10.1101/2023.09.20.558726. bioRxiv. 2023. Update in: Nat Cell Biol. 2024 Feb;26(2):250-262. doi: 10.1038/s41556-023-01337-z. PMID: 37790430 Free PMC article. Updated. Preprint.
Similar articles
-
Epithelial zonation along the mouse and human small intestine defines five discrete metabolic domains.bioRxiv [Preprint]. 2023 Sep 22:2023.09.20.558726. doi: 10.1101/2023.09.20.558726. bioRxiv. 2023. Update in: Nat Cell Biol. 2024 Feb;26(2):250-262. doi: 10.1038/s41556-023-01337-z. PMID: 37790430 Free PMC article. Updated. Preprint.
-
Changes in morphology and miRNAs expression in small intestines of Shaoxing ducks in response to high temperature.Mol Biol Rep. 2019 Aug;46(4):3843-3856. doi: 10.1007/s11033-019-04827-2. Epub 2019 May 2. Mol Biol Rep. 2019. PMID: 31049835
-
Regional epithelial cell diversity in the small intestine of pigs.J Anim Sci. 2023 Jan 3;101:skac318. doi: 10.1093/jas/skac318. J Anim Sci. 2023. PMID: 36183288 Free PMC article.
-
Absorption and metabolism of purines by the small intestine of the chicken.Comp Biochem Physiol A Comp Physiol. 1991;99(1-2):235-40. doi: 10.1016/0300-9629(91)90265-e. Comp Biochem Physiol A Comp Physiol. 1991. PMID: 1675952
-
Factors that influence absorption and secretion of calcium in the small intestine and colon.Am J Physiol. 1985 Feb;248(2 Pt 1):G147-57. doi: 10.1152/ajpgi.1985.248.2.G147. Am J Physiol. 1985. PMID: 3881991 Review.
Cited by
-
Tuft cells in the intestine, immunity and beyond.Nat Rev Gastroenterol Hepatol. 2024 Dec;21(12):852-868. doi: 10.1038/s41575-024-00978-1. Epub 2024 Sep 26. Nat Rev Gastroenterol Hepatol. 2024. PMID: 39327439 Review.
-
Spatially restricted immune and microbiota-driven adaptation of the gut.Nature. 2024 Dec;636(8042):447-456. doi: 10.1038/s41586-024-08216-z. Epub 2024 Nov 20. Nature. 2024. PMID: 39567686 Free PMC article.
-
Harnessing 3D models to uncover the mechanisms driving infectious and inflammatory disease in the intestine.BMC Biol. 2024 Dec 31;22(1):300. doi: 10.1186/s12915-024-02092-9. BMC Biol. 2024. PMID: 39736603 Free PMC article. Review.
-
Functional Diversity of Memory CD8 T Cells is Spatiotemporally Imprinted.bioRxiv [Preprint]. 2024 Mar 25:2024.03.20.585130. doi: 10.1101/2024.03.20.585130. bioRxiv. 2024. Update in: Nature. 2025 Mar;639(8054):483-492. doi: 10.1038/s41586-024-08466-x. PMID: 38585842 Free PMC article. Updated. Preprint.
References
-
- Brown H & Esterhazy D Intestinal immune compartmentalization: implications of tissue specific determinants in health and disease. Mucosal Immunol. 14, 1259–1270 (2021). - PubMed
-
- Altmann GG & Leblond CP Factors influencing villus size in the small intestine of adult rats as revealed by transposition of intestinal segments. Am. J. Anat 127, 15–36 (1970). - PubMed
-
- Bates MD et al. Novel genes and functional relationships in the adult mouse gastrointestinal tract identified by microarray analysis. Gastroenterology 122, 1467–1482 (2002). - PubMed
