

Oxygenation influences xylose fermentation and gene expression in the yeast genera Spathaspora and Scheffersomyces
- PMID: 38321504
- PMCID: PMC10848558
- DOI: 10.1186/s13068-024-02467-8
Oxygenation influences xylose fermentation and gene expression in the yeast genera Spathaspora and Scheffersomyces
Abstract
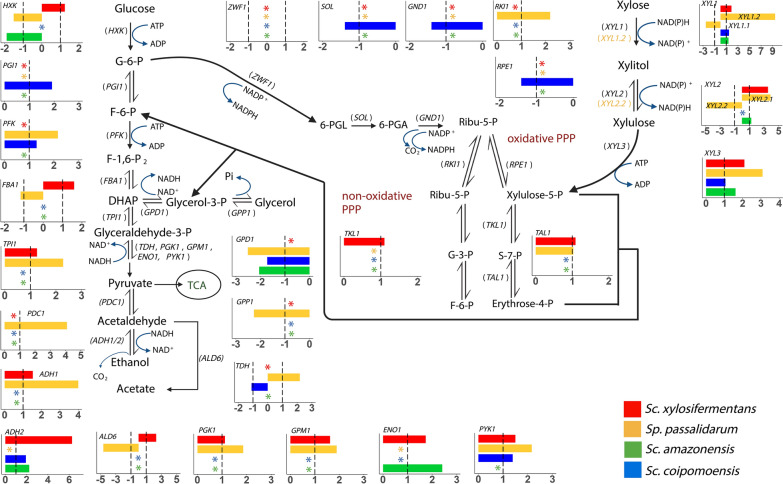
Background: Cost-effective production of biofuels from lignocellulose requires the fermentation of D-xylose. Many yeast species within and closely related to the genera Spathaspora and Scheffersomyces (both of the order Serinales) natively assimilate and ferment xylose. Other species consume xylose inefficiently, leading to extracellular accumulation of xylitol. Xylitol excretion is thought to be due to the different cofactor requirements of the first two steps of xylose metabolism. Xylose reductase (XR) generally uses NADPH to reduce xylose to xylitol, while xylitol dehydrogenase (XDH) generally uses NAD+ to oxidize xylitol to xylulose, creating an imbalanced redox pathway. This imbalance is thought to be particularly consequential in hypoxic or anoxic environments.
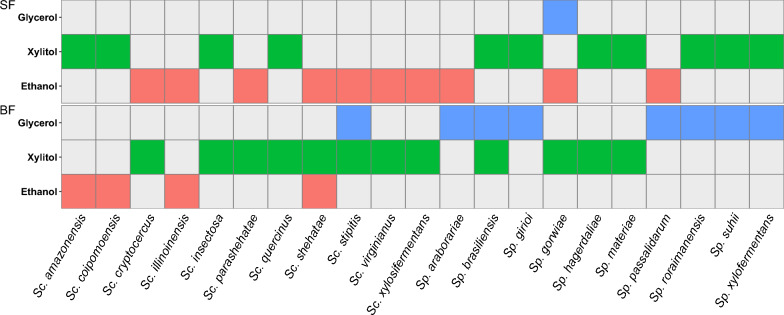
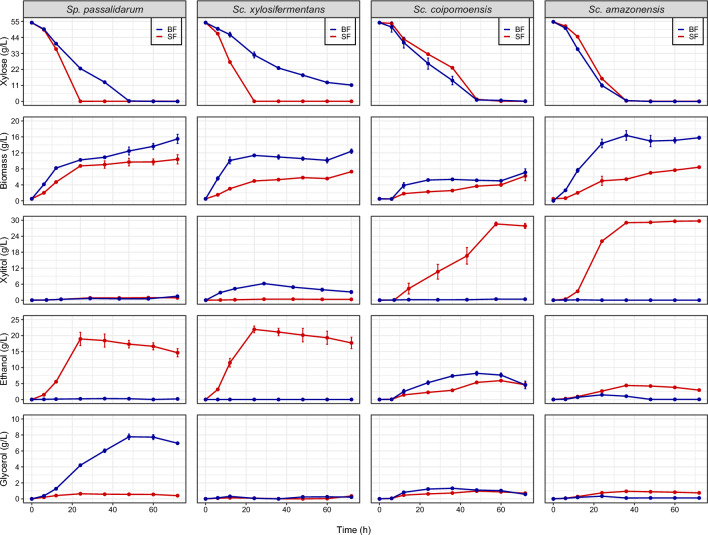
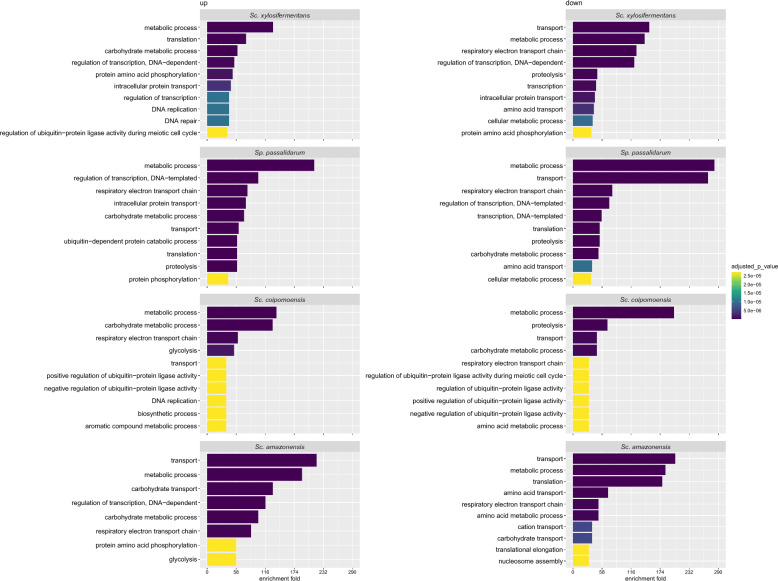
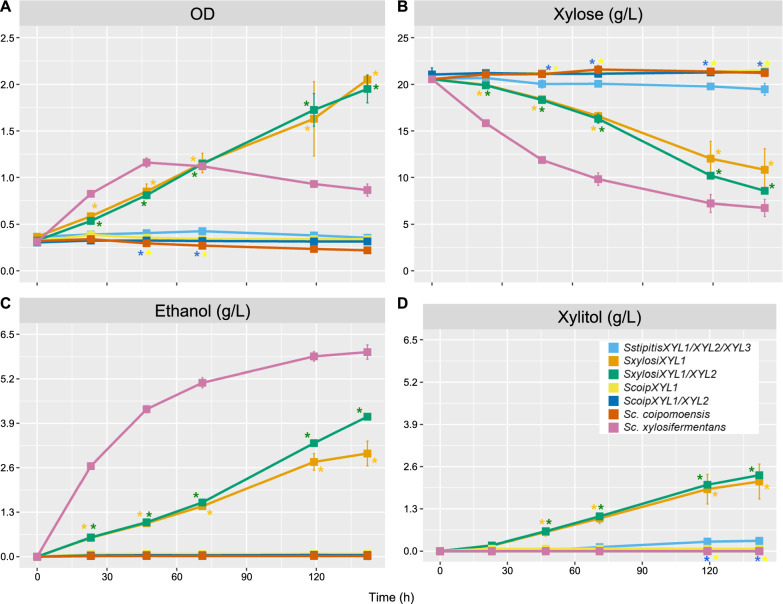
Results: We screened the growth of xylose-fermenting yeast species in high and moderate aeration and identified both ethanol producers and xylitol producers. Selected species were further characterized for their XR and XDH cofactor preferences by enzyme assays and gene expression patterns by RNA-Seq. Our data revealed that xylose metabolism is more redox balanced in some species, but it is strongly affected by oxygen levels. Under high aeration, most species switched from ethanol production to xylitol accumulation, despite the availability of ample oxygen to accept electrons from NADH. This switch was followed by decreases in enzyme activity and the expression of genes related to xylose metabolism, suggesting that bottlenecks in xylose fermentation are not always due to cofactor preferences. Finally, we expressed XYL genes from multiple Scheffersomyces species in a strain of Saccharomyces cerevisiae. Recombinant S. cerevisiae expressing XYL1 from Scheffersomyces xylosifermentans, which encodes an XR without a cofactor preference, showed improved anaerobic growth on xylose as the primary carbon source compared to S. cerevisiae strain expressing XYL genes from Scheffersomyces stipitis.
Conclusion: Collectively, our data do not support the hypothesis that xylitol accumulation occurs primarily due to differences in cofactor preferences between xylose reductase and xylitol dehydrogenase; instead, gene expression plays a major role in response to oxygen levels. We have also identified the yeast Sc. xylosifermentans as a potential source for genes that can be engineered into S. cerevisiae to improve xylose fermentation and biofuel production.
Keywords: Aeration; Cofactors; Ethanol; Gene expression; Serinales (CUG-Ser1 clade); Xylitol; Xylitol dehydrogenase; Xylose fermentation; Xylose reductase.
© 2024. The Author(s).
Conflict of interest statement
The Wisconsin Alumni Research Foundation has filed a provisional patent application based on the contents of this manuscript with KOB, TKS, and CTH as inventors.
Figures

Similar articles
-
Exploring xylose metabolism in Spathaspora species: XYL1.2 from Spathaspora passalidarum as the key for efficient anaerobic xylose fermentation in metabolic engineered Saccharomyces cerevisiae.Biotechnol Biofuels. 2016 Aug 5;9:167. doi: 10.1186/s13068-016-0570-6. eCollection 2016. Biotechnol Biofuels. 2016. PMID: 27499810 Free PMC article.
-
Enhanced xylose fermentation by engineered yeast expressing NADH oxidase through high cell density inoculums.J Ind Microbiol Biotechnol. 2017 Mar;44(3):387-395. doi: 10.1007/s10295-016-1899-3. Epub 2017 Jan 9. J Ind Microbiol Biotechnol. 2017. PMID: 28070721
-
A genetic overhaul of Saccharomyces cerevisiae 424A(LNH-ST) to improve xylose fermentation.J Ind Microbiol Biotechnol. 2011 May;38(5):617-26. doi: 10.1007/s10295-010-0806-6. Epub 2010 Aug 17. J Ind Microbiol Biotechnol. 2011. PMID: 20714780
-
Deletion of FPS1, encoding aquaglyceroporin Fps1p, improves xylose fermentation by engineered Saccharomyces cerevisiae.Appl Environ Microbiol. 2013 May;79(10):3193-201. doi: 10.1128/AEM.00490-13. Epub 2013 Mar 8. Appl Environ Microbiol. 2013. PMID: 23475614 Free PMC article.
-
Metabolic engineering for improved fermentation of pentoses by yeasts.Appl Microbiol Biotechnol. 2004 Feb;63(5):495-509. doi: 10.1007/s00253-003-1450-0. Epub 2003 Nov 1. Appl Microbiol Biotechnol. 2004. PMID: 14595523 Review.
Cited by
-
Exploring xylose metabolism in non-conventional yeasts: kinetic characterization and product accumulation under different aeration conditions.J Ind Microbiol Biotechnol. 2024 Jan 9;51:kuae023. doi: 10.1093/jimb/kuae023. J Ind Microbiol Biotechnol. 2024. PMID: 38936832 Free PMC article.
-
Engineering transcriptional regulatory networks for improving second-generation fuel ethanol production in Saccharomyces cerevisiae.Synth Syst Biotechnol. 2024 Oct 28;10(1):207-217. doi: 10.1016/j.synbio.2024.10.006. eCollection 2025. Synth Syst Biotechnol. 2024. PMID: 39558946 Free PMC article. Review.
-
Genomic characteristics and genetic manipulation of the marine yeast Scheffersomyces spartinae.Appl Microbiol Biotechnol. 2024 Dec 19;108(1):539. doi: 10.1007/s00253-024-13382-1. Appl Microbiol Biotechnol. 2024. PMID: 39702830 Free PMC article.
References
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous