

Interface Gain-of-Function Mutations in TLR7 Cause Systemic and Neuro-inflammatory Disease
- PMID: 38324161
- PMCID: PMC10850255
- DOI: 10.1007/s10875-024-01660-6
Interface Gain-of-Function Mutations in TLR7 Cause Systemic and Neuro-inflammatory Disease
Abstract
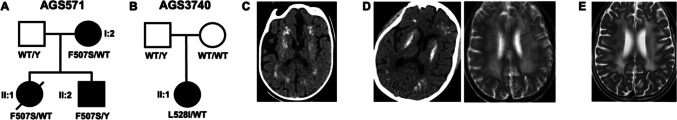
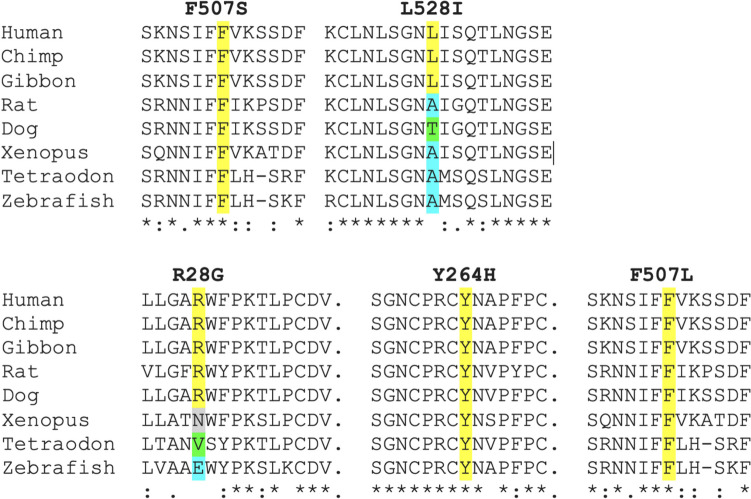
TLR7 recognizes pathogen-derived single-stranded RNA (ssRNA), a function integral to the innate immune response to viral infection. Notably, TLR7 can also recognize self-derived ssRNA, with gain-of-function mutations in human TLR7 recently identified to cause both early-onset systemic lupus erythematosus (SLE) and neuromyelitis optica. Here, we describe two novel mutations in TLR7, F507S and L528I. While the L528I substitution arose de novo, the F507S mutation was present in three individuals from the same family, including a severely affected male, notably given that the TLR7 gene is situated on the X chromosome and that all other cases so far described have been female. The observation of mutations at residues 507 and 528 of TLR7 indicates the importance of the TLR7 dimerization interface in maintaining immune homeostasis, where we predict that altered homo-dimerization enhances TLR7 signaling. Finally, while mutations in TLR7 can result in SLE-like disease, our data suggest a broader phenotypic spectrum associated with TLR7 gain-of-function, including significant neurological involvement.
Keywords: TLR7; stem cell transplantation; systemic lupus erythematosus.
© 2024. The Author(s).
Conflict of interest statement
Y.J.C. is an associate editor of the Journal of Clinical Immunology. The authors declare no other conflicts of interest, financial or otherwise.
Figures


References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
