

Itaconate impairs immune control of Plasmodium by enhancing mtDNA-mediated PD-L1 expression in monocyte-derived dendritic cells
- PMID: 38325373
- PMCID: PMC10940217
- DOI: 10.1016/j.cmet.2024.01.008
Itaconate impairs immune control of Plasmodium by enhancing mtDNA-mediated PD-L1 expression in monocyte-derived dendritic cells
Abstract
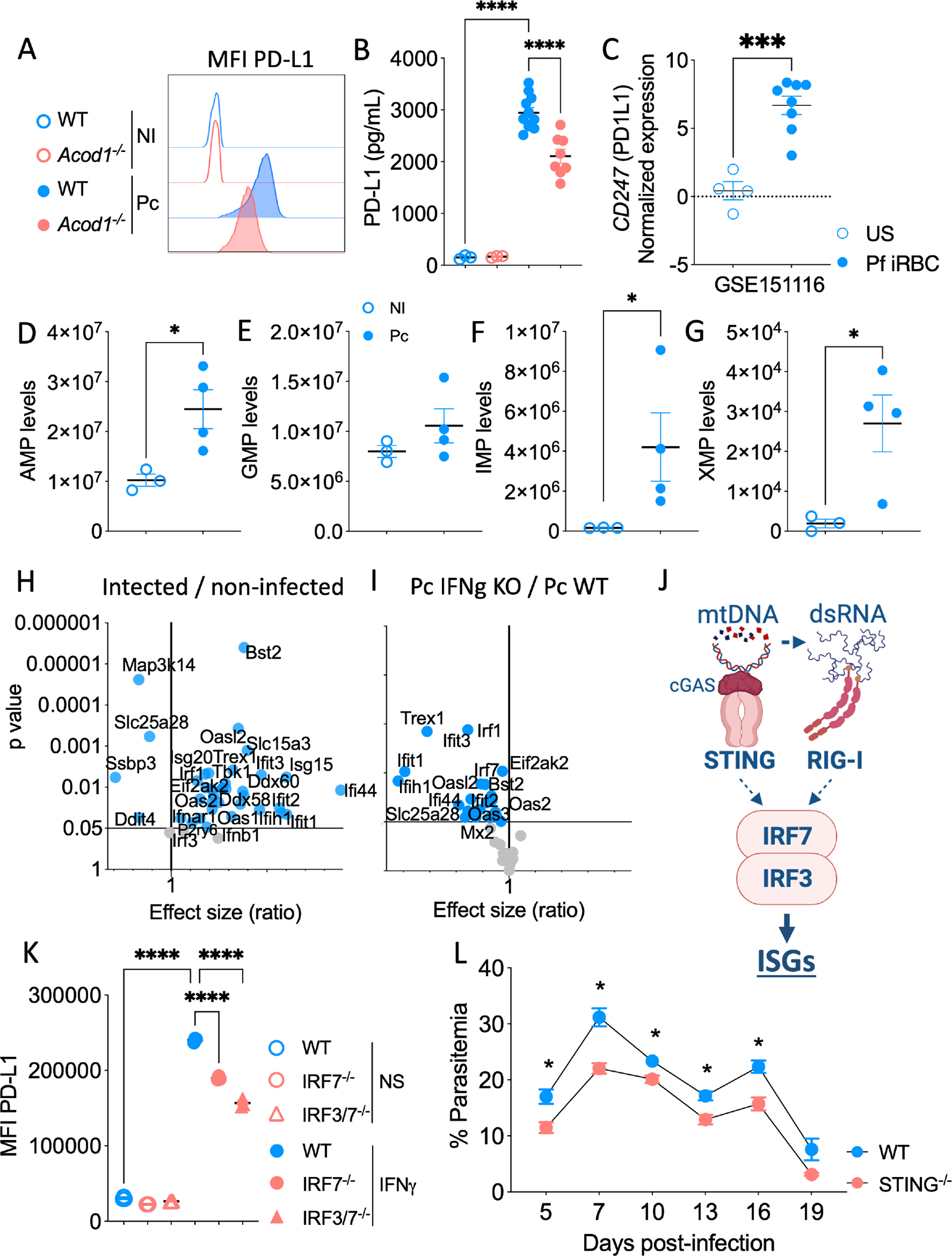
Severe forms of malaria are associated with systemic inflammation and host metabolism disorders; however, the interplay between these outcomes is poorly understood. Using a Plasmodium chabaudi model of malaria, we demonstrate that interferon (IFN) γ boosts glycolysis in splenic monocyte-derived dendritic cells (MODCs), leading to itaconate accumulation and disruption in the TCA cycle. Increased itaconate levels reduce mitochondrial functionality, which associates with organellar nucleic acid release and MODC restraint. We hypothesize that dysfunctional mitochondria release degraded DNA into the cytosol. Once mitochondrial DNA is sensitized, the activation of IRF3 and IRF7 promotes the expression of IFN-stimulated genes and checkpoint markers. Indeed, depletion of the STING-IRF3/IRF7 axis reduces PD-L1 expression, enabling activation of CD8+ T cells that control parasite proliferation. In summary, mitochondrial disruption caused by itaconate in MODCs leads to a suppressive effect in CD8+ T cells, which enhances parasitemia. We provide evidence that ACOD1 and itaconate are potential targets for adjunct antimalarial therapy.
Keywords: PD-1; PD-L1; Plasmodium chabaudi; TCA cycle; cGAS-STING; immuno checkpoint markers; inate immunity; itaconate; itaconic acid; lymphocytes; malaria; metabolism; methylenesuccinic acid; mitochondria; mitochondrial DNA; monocyte-derived dendritic cells; mtDNA.
Published by Elsevier Inc.
Conflict of interest statement
Declaration of interests The authors declare no competing interests.
Figures






Comment in
-
Itaconate boosts malaria via induction of PD-L1.Cell Metab. 2024 Mar 5;36(3):457-458. doi: 10.1016/j.cmet.2024.02.005. Cell Metab. 2024. PMID: 38447526
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Research Materials
