

High-efficiency genome editing by Cas12a ribonucleoprotein complex in Euglena gracilis
- PMID: 38332568
- PMCID: PMC10884871
- DOI: 10.1111/1751-7915.14393
High-efficiency genome editing by Cas12a ribonucleoprotein complex in Euglena gracilis
Abstract
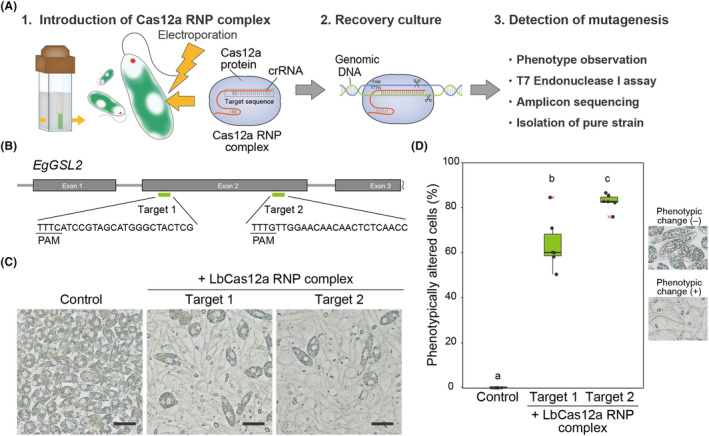
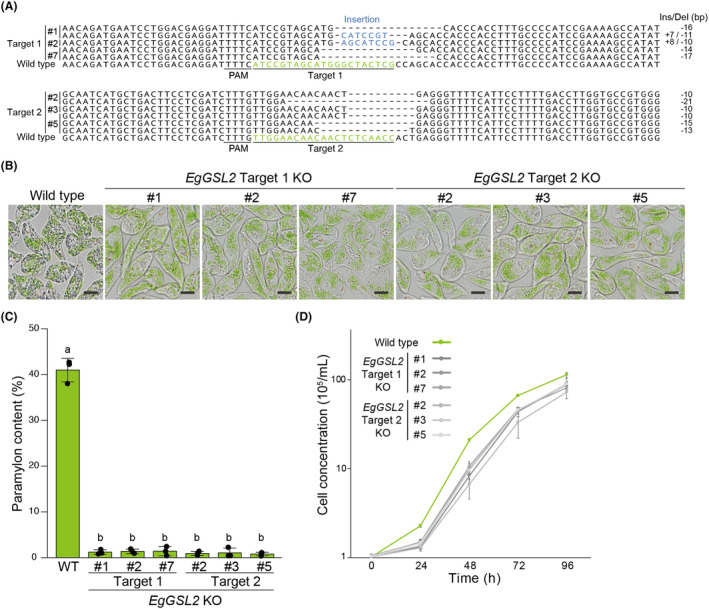
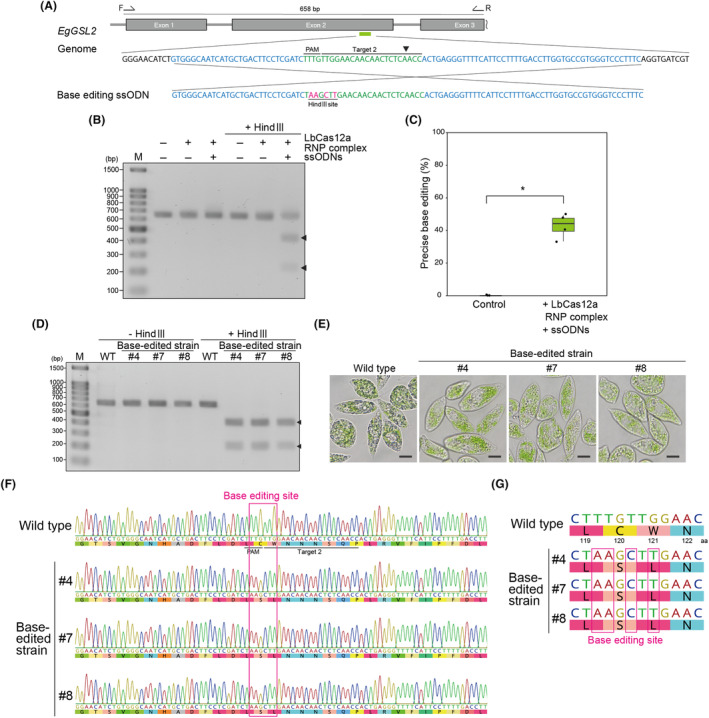
Transgene-free genome editing based on clustered regularly interspaced short palindromic repeats (CRISPR) technology is key to achieving genetic engineering in microalgae for basic research and industrial applications. Euglena gracilis, a unicellular phytoflagellate microalga, is a promising biomaterial for foods, feeds, cosmetics and biofuels. However, methods for the genetic manipulation of E. gracilis are still limited. Here, we developed a high-efficiency, transgene-free genome editing method for E. gracilis using Lachnospiraceae bacterium CRISPR-associated protein 12a (LbCas12a) ribonucleoprotein (RNP) complex, which complements the previously established Cas9 RNP-based method. Through the direct delivery of LbCas12a-containing RNPs, our method reached mutagenesis rates of approximately 77.2-94.5% at two different E. gracilis target genes, Glucan synthase-like 2 (EgGSL2) and a phytoene synthase gene (EgcrtB). Moreover, in addition to targeted mutagenesis, we demonstrated efficient knock-in and base editing at the target site using LbCas12a-based RNPs with a single-stranded DNA donor template in E. gracilis. This study extends the genetic engineering capabilities of Euglena to accelerate its basic use for research and engineering for bioproduction.
© 2024 The Authors. Microbial Biotechnology published by Applied Microbiology International and John Wiley & Sons Ltd.
Conflict of interest statement
This study was partially supported by a matching fund‐based research program between RIKEN and Euglena Co., Ltd.
Figures



Similar articles
-
Highly Efficient CRISPR-Associated Protein 9 Ribonucleoprotein-Based Genome Editing in Euglena gracilis.STAR Protoc. 2020 Jun 3;1(1):100023. doi: 10.1016/j.xpro.2020.100023. eCollection 2020 Jun 19. STAR Protoc. 2020. PMID: 33111076 Free PMC article.
-
Highly efficient transgene-free targeted mutagenesis and single-stranded oligodeoxynucleotide-mediated precise knock-in in the industrial microalga Euglena gracilis using Cas9 ribonucleoproteins.Plant Biotechnol J. 2019 Nov;17(11):2032-2034. doi: 10.1111/pbi.13174. Epub 2019 Jun 11. Plant Biotechnol J. 2019. PMID: 31131534 Free PMC article. No abstract available.
-
Delivery of CRISPR-Cas12a Ribonucleoprotein Complex for Genome Editing in an Embryogenic Citrus Cell Line.Methods Mol Biol. 2023;2653:153-171. doi: 10.1007/978-1-0716-3131-7_10. Methods Mol Biol. 2023. PMID: 36995625
-
Highly Efficient Genome Editing in Plant Protoplasts by Ribonucleoprotein Delivery of CRISPR-Cas12a Nucleases.Front Genome Ed. 2022 Jan 31;4:780238. doi: 10.3389/fgeed.2022.780238. eCollection 2022. Front Genome Ed. 2022. PMID: 35174354 Free PMC article. Review.
-
CRISPR ribonucleoprotein-mediated genetic engineering in plants.Plant Commun. 2021 Feb 10;2(2):100168. doi: 10.1016/j.xplc.2021.100168. eCollection 2021 Mar 8. Plant Commun. 2021. PMID: 33898980 Free PMC article. Review.
Cited by
-
Changes in Gravitaxis and Gene-Expression in an Euglena gracilis Culture over Time.Biomolecules. 2024 Mar 9;14(3):327. doi: 10.3390/biom14030327. Biomolecules. 2024. PMID: 38540747 Free PMC article.
References
-
- Abràmoff, M.D. , Magalhães, P.J. & Ram, S.J. (2004) Image processing with ImageJ. Biophotonics International, 11, 36–42.
Publication types
MeSH terms
Substances
Grants and funding
- 21K05511/Japan Science and Technology Agency Grant-in-Aid for Scientific Research (C)
- JPMJOP1832/Program on Open Innovation Platform with Enterprises, Research Institute and Academia
- JPMJSA2204/Science and Technology Research Partnership for Sustainable Development
- JPMJGX23B0/Green Technologies for Excellence Program
LinkOut - more resources
Full Text Sources
