

ALS-related p97 R155H mutation disrupts lysophagy in iPSC-derived motor neurons
- PMID: 38335961
- PMCID: PMC10937112
- DOI: 10.1016/j.stemcr.2024.01.002
ALS-related p97 R155H mutation disrupts lysophagy in iPSC-derived motor neurons
Erratum in
-
ALS-related p97 R155H mutation disrupts lysophagy in iPSC-derived motor neurons.Stem Cell Reports. 2025 May 13;20(5):102478. doi: 10.1016/j.stemcr.2025.102478. Epub 2025 Mar 28. Stem Cell Reports. 2025. PMID: 40157354 Free PMC article. No abstract available.
Abstract
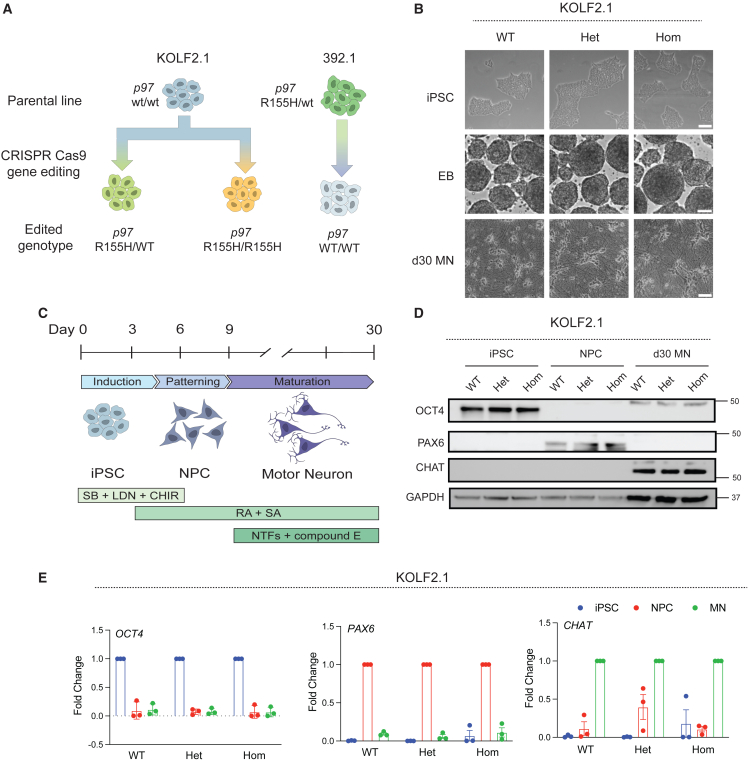
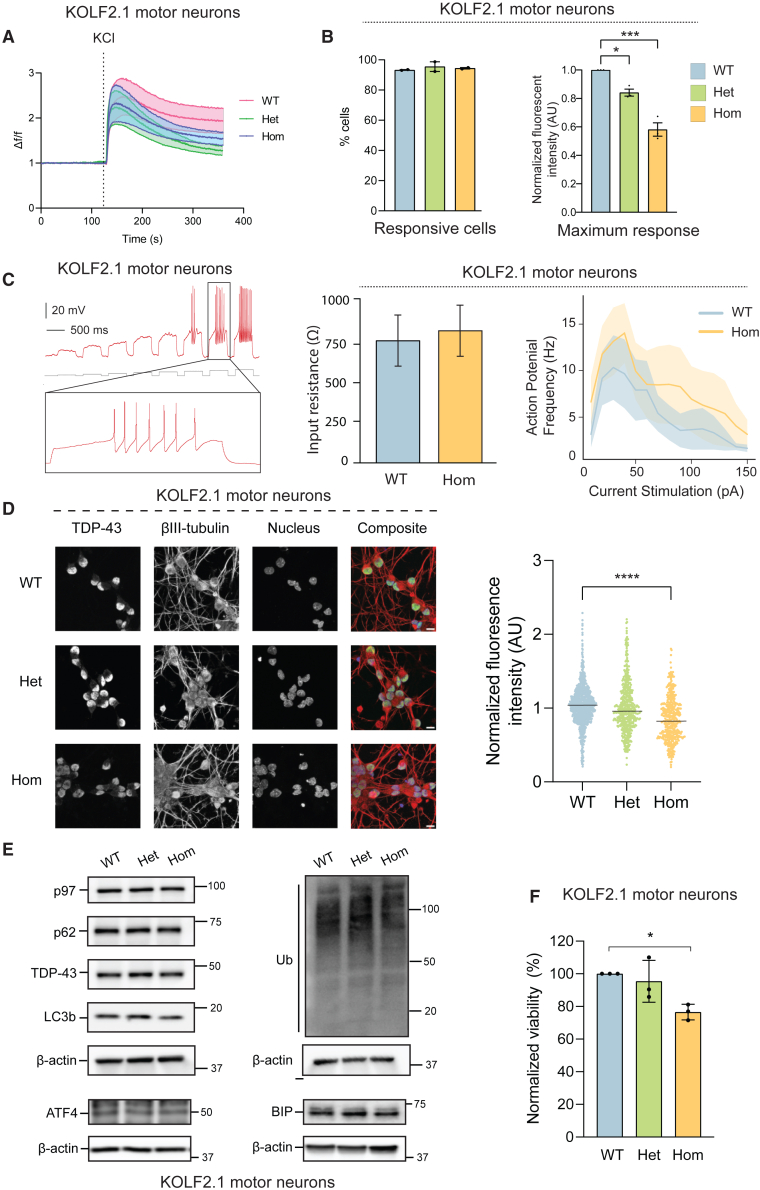
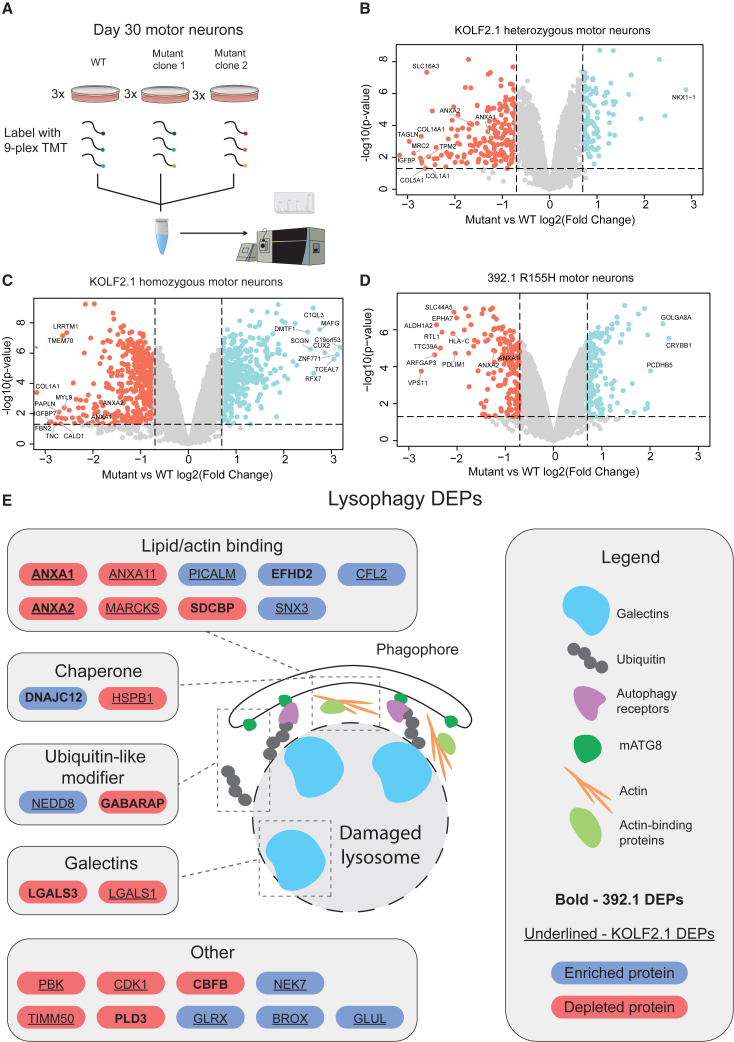
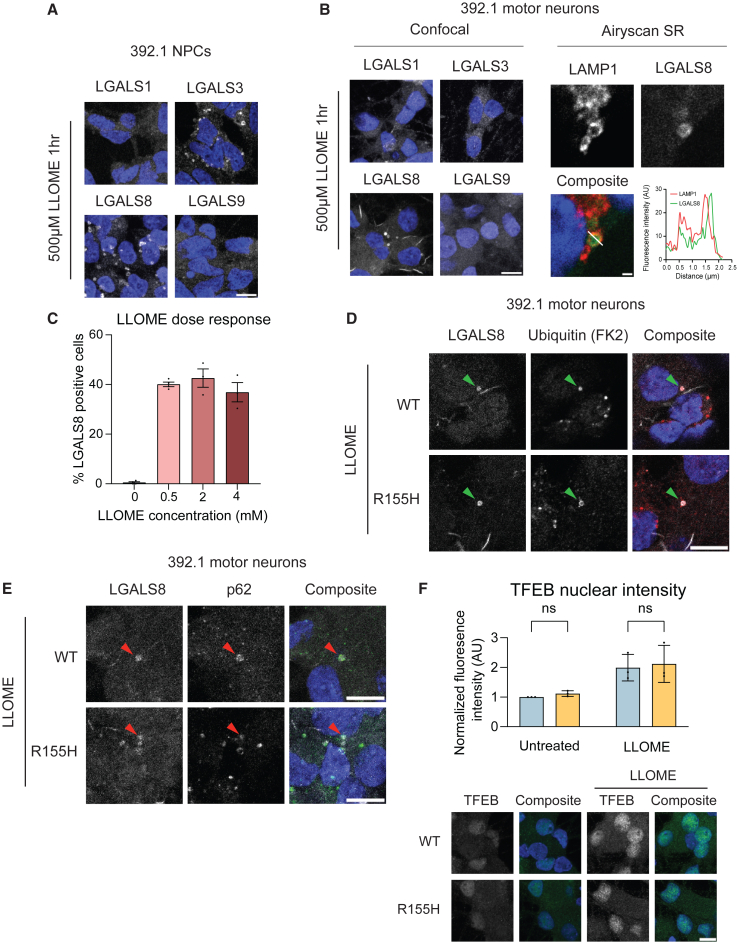
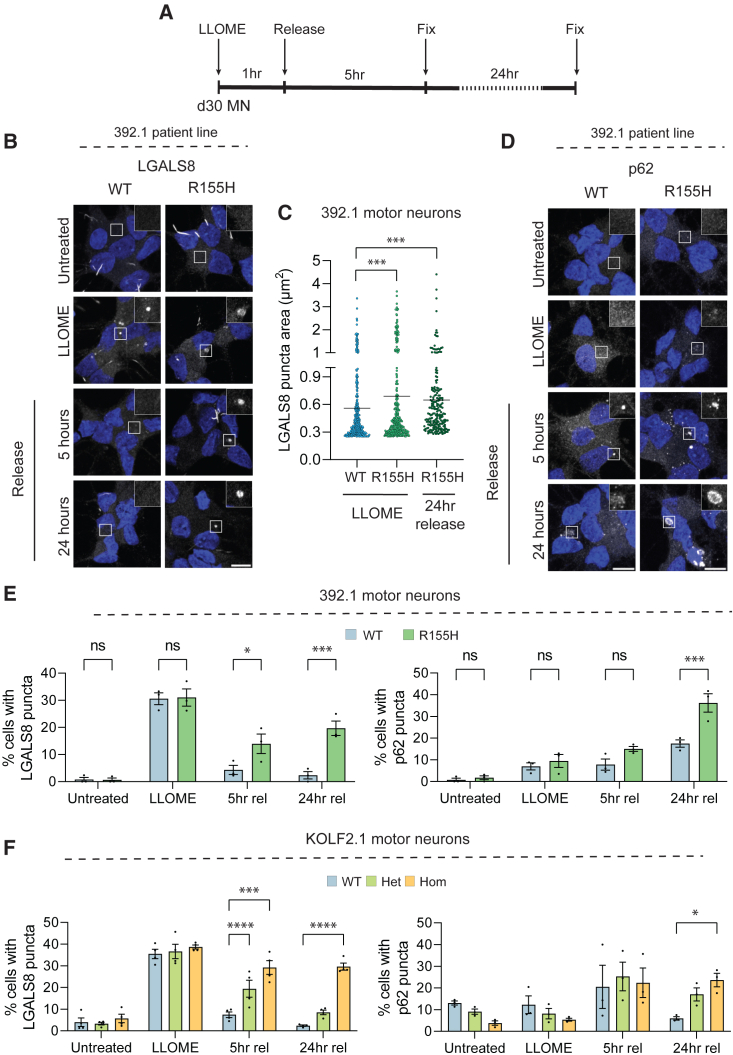
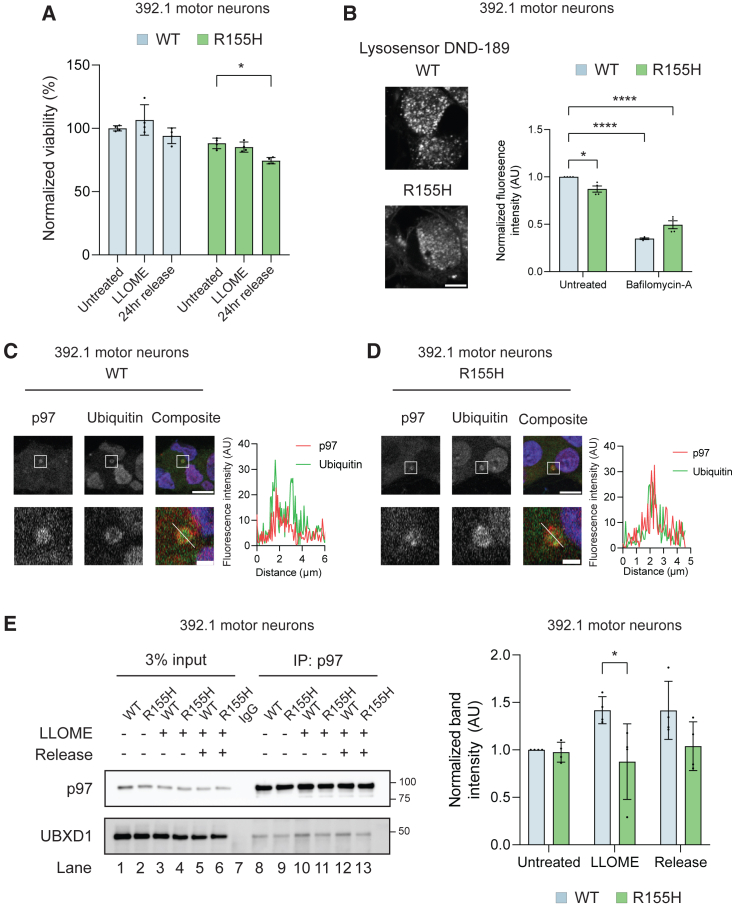
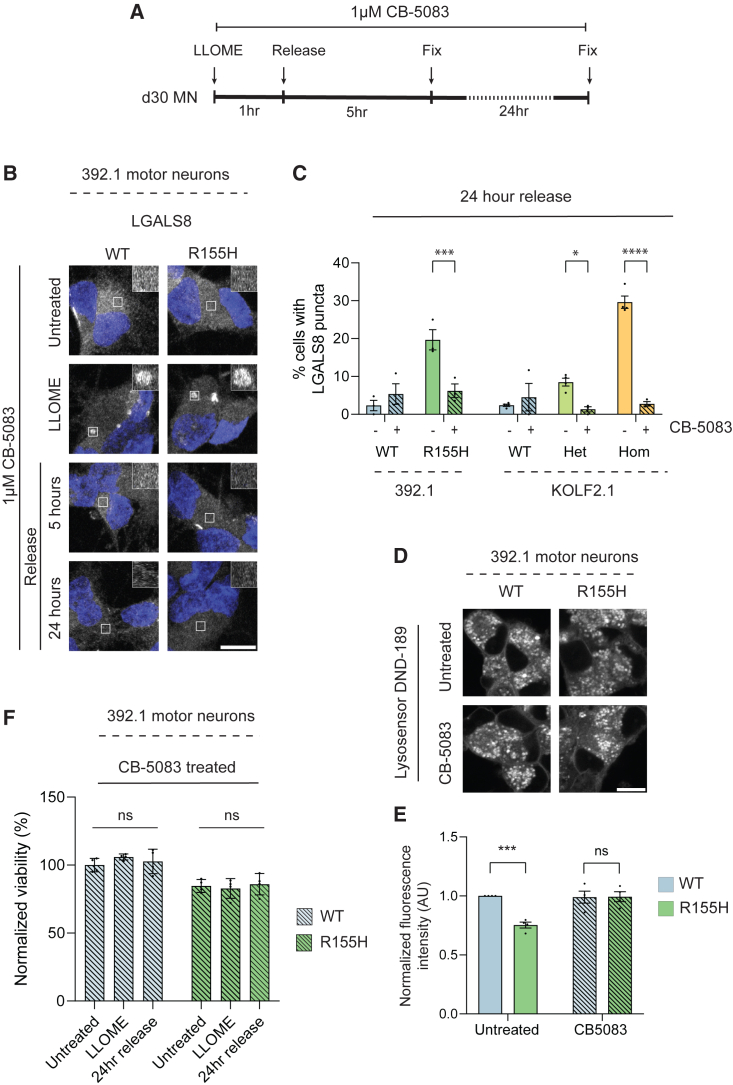
Mutations in the AAA+ ATPase p97 cause multisystem proteinopathy 1, which includes amyotrophic lateral sclerosis; however, the pathogenic mechanisms that contribute to motor neuron loss remain obscure. Here, we use two induced pluripotent stem cell models differentiated into spinal motor neurons to investigate how p97 mutations perturb the motor neuron proteome. Using quantitative proteomics, we find that motor neurons harboring the p97 R155H mutation have deficits in the selective autophagy of lysosomes (lysophagy). p97 R155H motor neurons are unable to clear damaged lysosomes and have reduced viability. Lysosomes in mutant motor neurons have increased pH compared with wild-type cells. The clearance of damaged lysosomes involves UBXD1-p97 interaction, which is disrupted in mutant motor neurons. Finally, inhibition of the ATPase activity of p97 using the inhibitor CB-5083 rescues lysophagy defects in mutant motor neurons. These results add to the evidence that endo-lysosomal dysfunction is a key aspect of disease pathogenesis in p97-related disorders.
Keywords: ALS; autophagy; galectin; lysophagy; lysosome; mitochondria; p97; proteomics.
Copyright © 2024 The Author(s). Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests The authors declare no competing interests.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
- R01 GM127557/GM/NIGMS NIH HHS/United States
- R01 NS102937/NS/NINDS NIH HHS/United States
- P50 MH122379/MH/NIMH NIH HHS/United States
- R01 GM067945/GM/NIGMS NIH HHS/United States
- R01 AA026256/AA/NIAAA NIH HHS/United States
- R01 MH128235/MH/NIMH NIH HHS/United States
- K24 AR073317/AR/NIAMS NIH HHS/United States
- R01 AG031867/AG/NIA NIH HHS/United States
- R21 NS123631/NS/NINDS NIH HHS/United States
- K12 GM133314/GM/NIGMS NIH HHS/United States
- R01 GM132129/GM/NIGMS NIH HHS/United States
- R01 NS105628/NS/NINDS NIH HHS/United States
LinkOut - more resources
Full Text Sources
Medical
Miscellaneous
