

Multiple Genes Core to ERAD, Macroautophagy and Lysosomal Degradation Pathways Participate in the Proteostasis Response in α1-Antitrypsin Deficiency
- PMID: 38336172
- PMCID: PMC11053228
- DOI: 10.1016/j.jcmgh.2024.02.006
Multiple Genes Core to ERAD, Macroautophagy and Lysosomal Degradation Pathways Participate in the Proteostasis Response in α1-Antitrypsin Deficiency
Abstract
Background & aims: In the classic form of α1-antitrypsin deficiency (ATD), the misfolded α1-antitrypsin Z (ATZ) variant accumulates in the endoplasmic reticulum (ER) of liver cells. A gain-of-function proteotoxic mechanism is responsible for chronic liver disease in a subgroup of homozygotes. Proteostatic response pathways, including conventional endoplasmic reticulum-associated degradation and autophagy, have been proposed as the mechanisms that allow cellular adaptation and presumably protection from the liver disease phenotype. Recent studies have concluded that a distinct lysosomal pathway called endoplasmic reticulum-to-lysosome completely supplants the role of the conventional macroautophagy pathway in degradation of ATZ. Here, we used several state-of-the-art approaches to characterize the proteostatic responses more fully in cellular systems that model ATD.
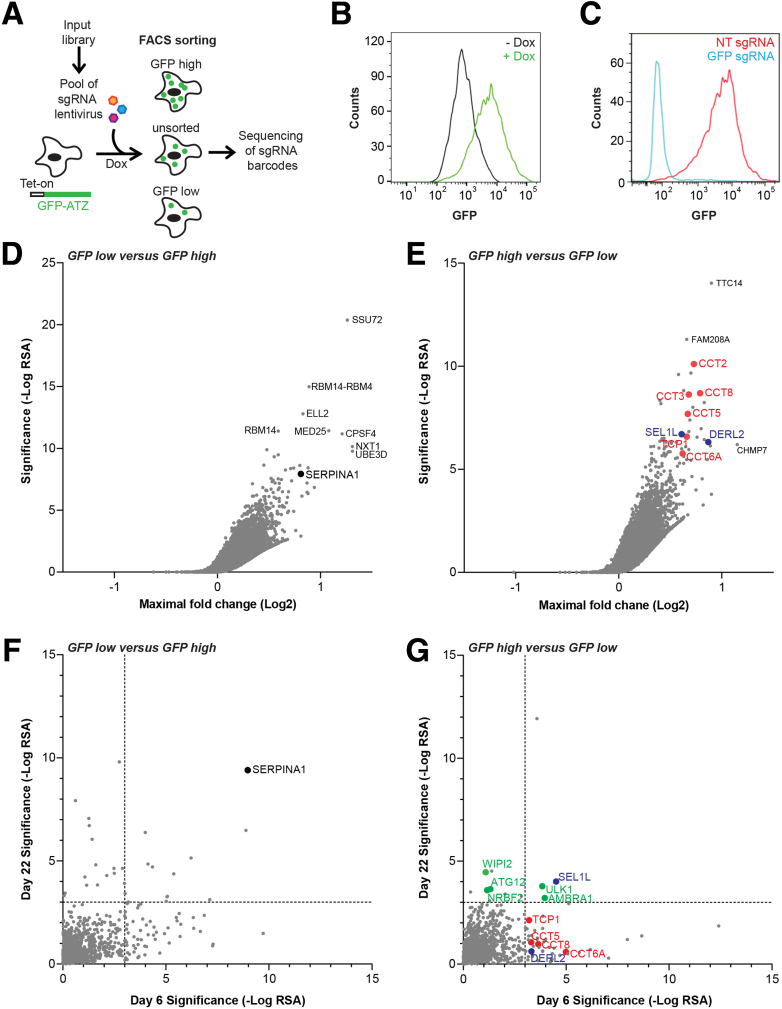
Methods: We used clustered regularly interspaced short palindromic repeats (CRISPR)-mediated genome editing coupled to a cell selection step by fluorescence-activated cell sorter to perform screening for proteostasis genes that regulate ATZ accumulation and combined that with selective genome editing in 2 other model systems.
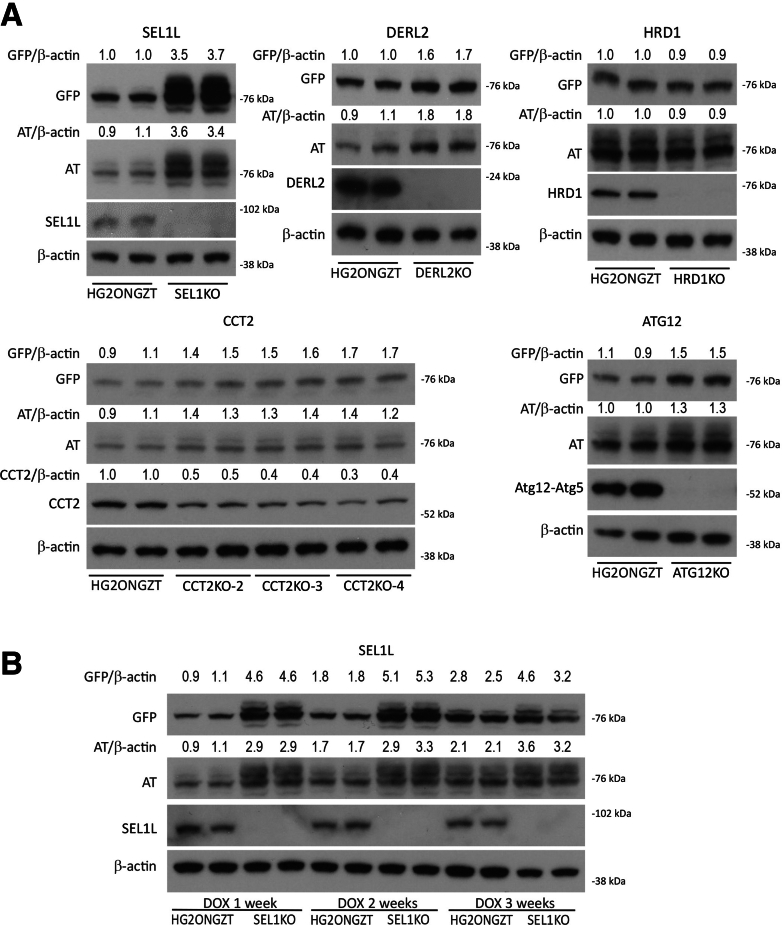
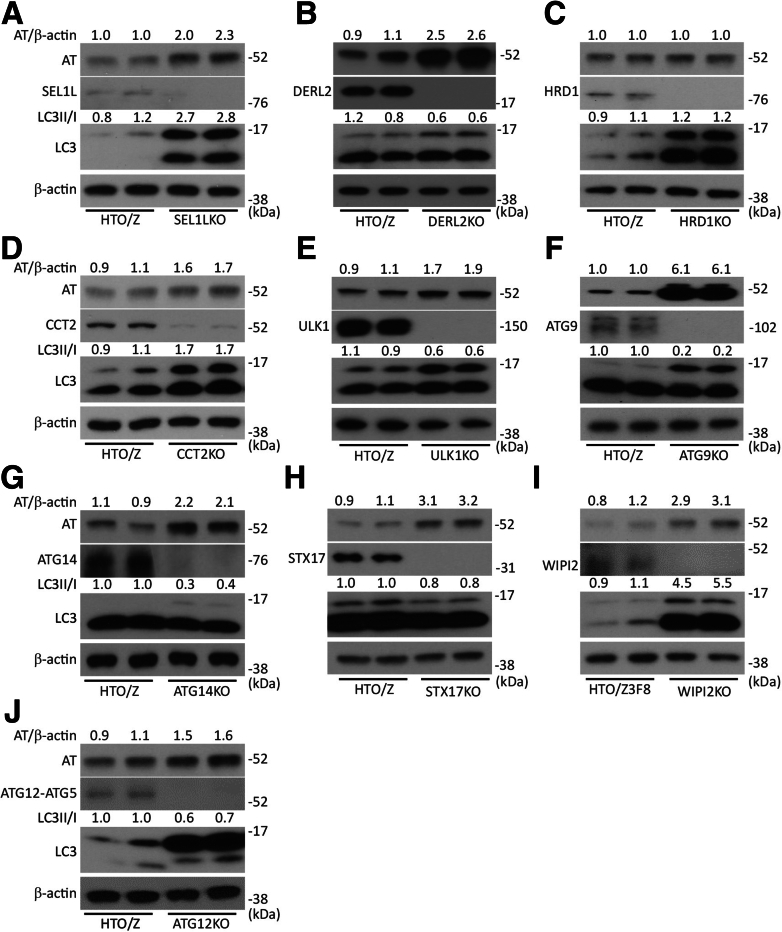
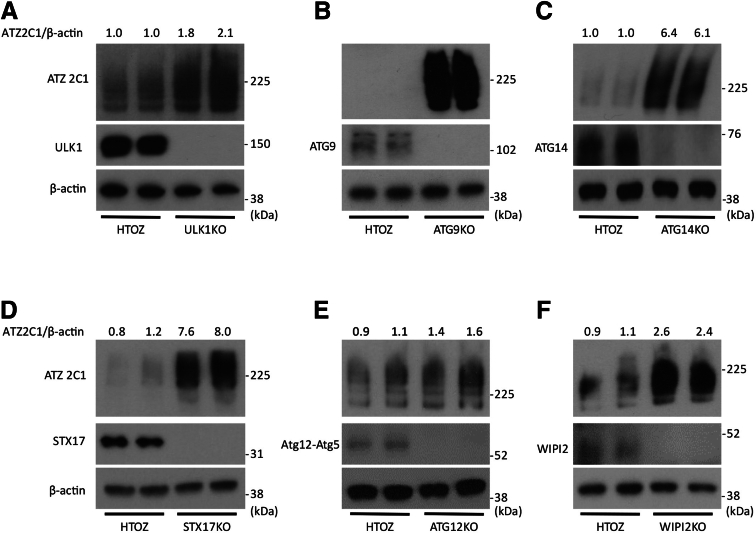
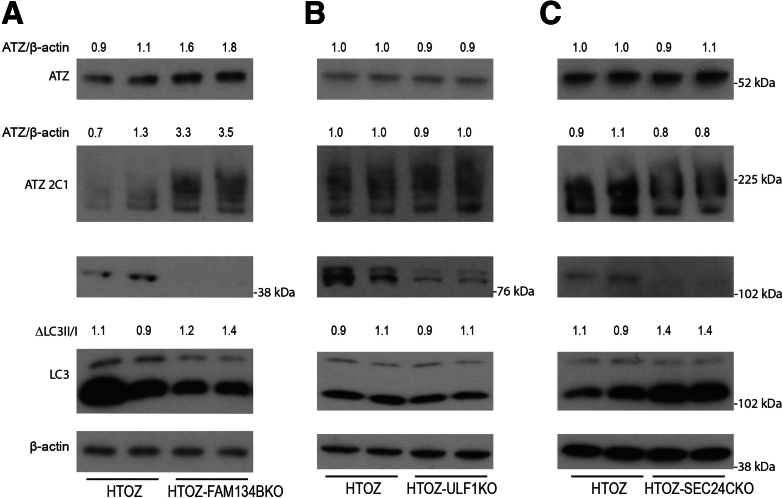
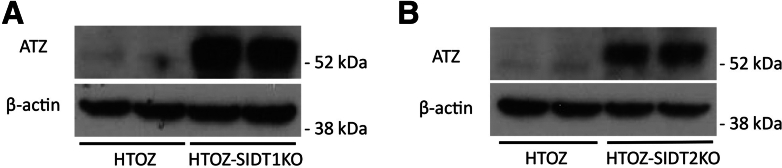
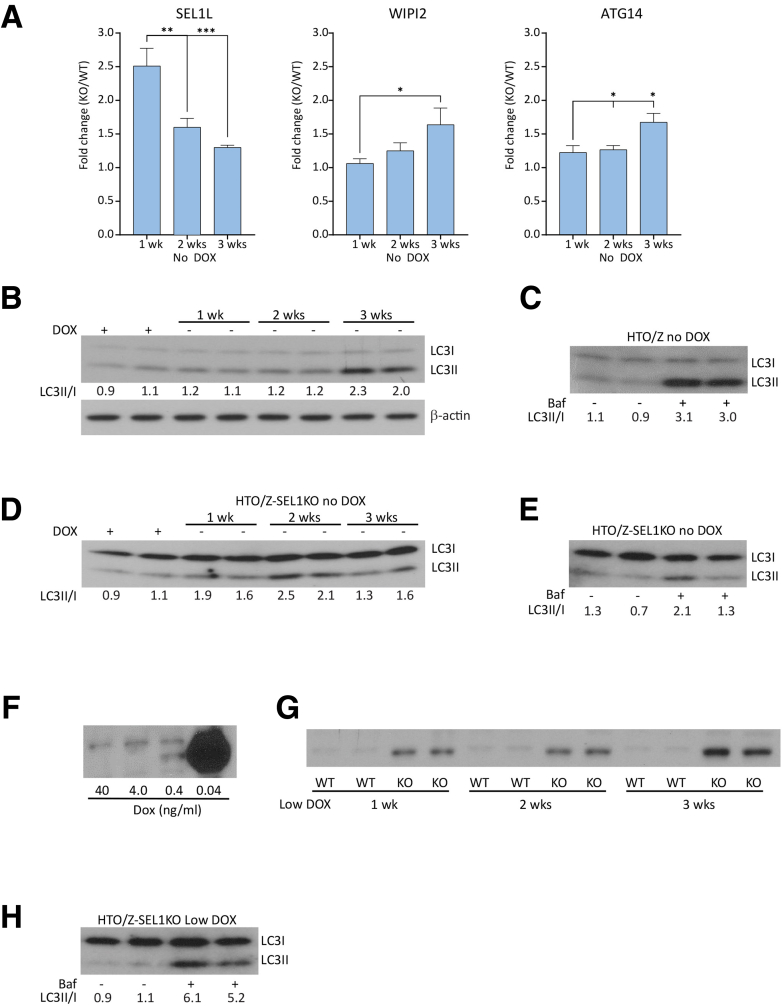
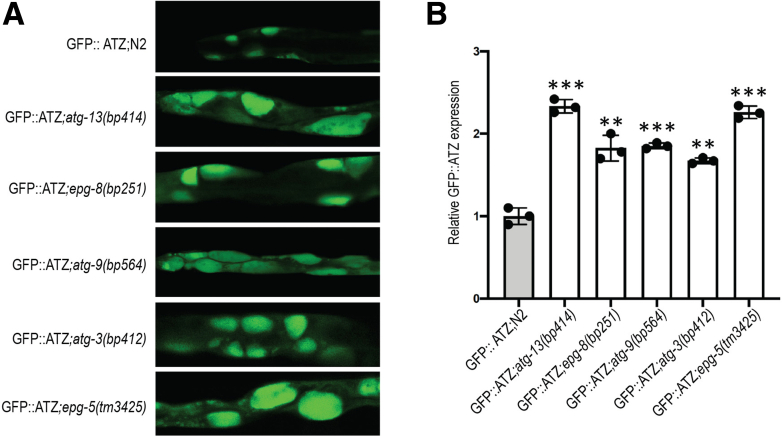
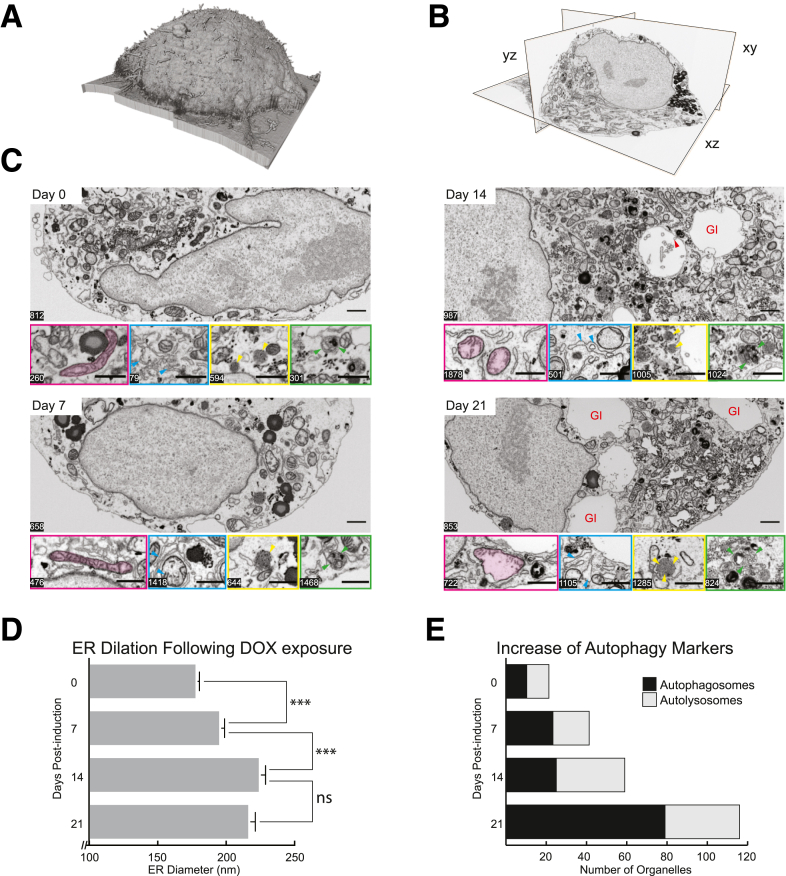
Results: Endoplasmic reticulum-associated degradation genes are key early regulators and multiple autophagy genes, from classic as well as from ER-to-lysosome and other newly described ER-phagy pathways, participate in degradation of ATZ in a manner that is temporally regulated and evolves as ATZ accumulation persists. Time-dependent changes in gene expression are accompanied by specific ultrastructural changes including dilation of the ER, formation of globular inclusions, budding of autophagic vesicles, and alterations in the overall shape and component parts of mitochondria.
Conclusions: Macroautophagy is a critical component of the proteostasis response to cellular ATZ accumulation and it becomes more important over time as ATZ synthesis continues unabated. Multiple subtypes of macroautophagy and nonautophagic lysosomal degradative pathways are needed to respond to the high concentrations of misfolded protein that characterizes ATD and these pathways are attractive candidates for genetic variants that predispose to the hepatic phenotype.
Keywords: Aggregation-Prone Proteins; Autophagy; Liver Disease; Proteasome; α1-Antitrypsin Deficiency.
Copyright © 2024 The Authors. Published by Elsevier Inc. All rights reserved.
Figures

References
-
- Khaminets A., Heinrich T., Mari M., et al. Regulation of endoplasmic reticulum turnover by selective autophagy. Nature. 2015;522:354–358. - PubMed
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Research Materials
