

Coevolution of RNA and protein subunits in RNase P and RNase MRP, two RNA processing enzymes
- PMID: 38336296
- PMCID: PMC10966300
- DOI: 10.1016/j.jbc.2024.105729
Coevolution of RNA and protein subunits in RNase P and RNase MRP, two RNA processing enzymes
Abstract
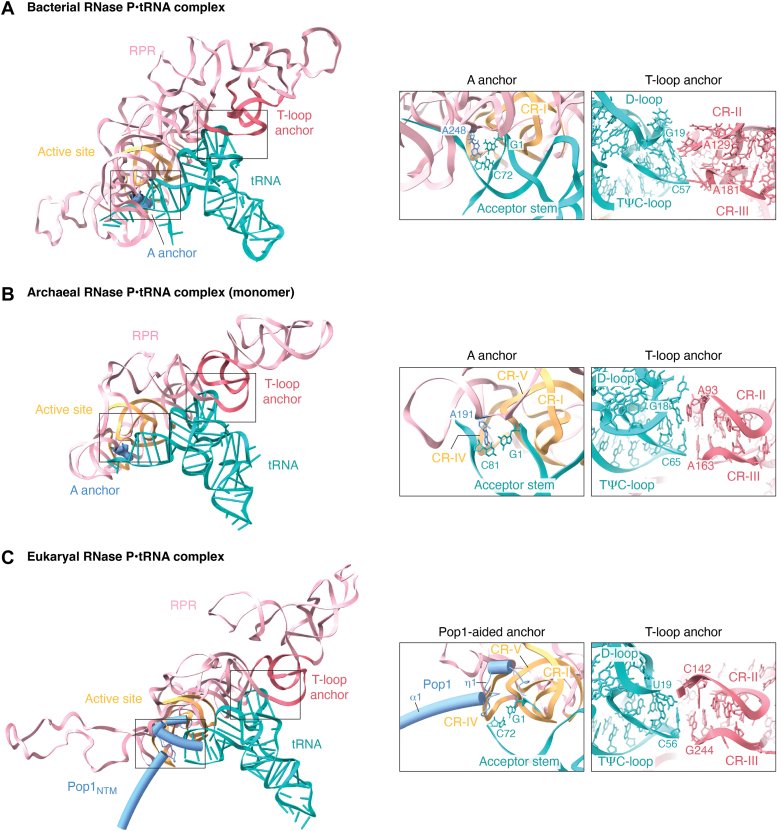
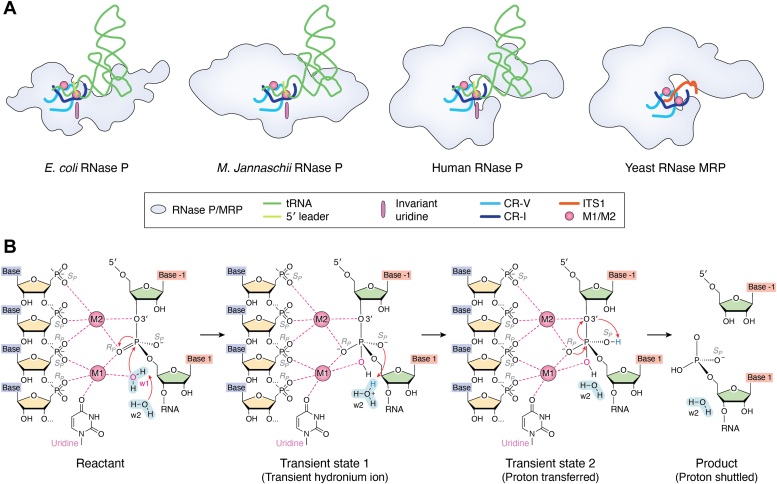
RNase P and RNase mitochondrial RNA processing (MRP) are ribonucleoproteins (RNPs) that consist of a catalytic RNA and a varying number of protein cofactors. RNase P is responsible for precursor tRNA maturation in all three domains of life, while RNase MRP, exclusive to eukaryotes, primarily functions in rRNA biogenesis. While eukaryotic RNase P is associated with more protein cofactors and has an RNA subunit with fewer auxiliary structural elements compared to its bacterial cousin, the double-anchor precursor tRNA recognition mechanism has remarkably been preserved during evolution. RNase MRP shares evolutionary and structural similarities with RNase P, preserving the catalytic core within the RNA moiety inherited from their common ancestor. By incorporating new protein cofactors and RNA elements, RNase MRP has established itself as a distinct RNP capable of processing ssRNA substrates. The structural information on RNase P and MRP helps build an evolutionary trajectory, depicting how emerging protein cofactors harmonize with the evolution of RNA to shape different functions for RNase P and MRP. Here, we outline the structural and functional relationship between RNase P and MRP to illustrate the coevolution of RNA and protein cofactors, a key driver for the extant, diverse RNP world.
Keywords: RNase MRP; RNase P; coevolution; cryo-EM; ribonucleoprotein.
Copyright © 2024 The Authors. Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Conflict of interest The authors declare that they have no conflicts of interest with the contents of this article.
Figures




Similar articles
-
Eukaryotic ribonuclease P: a plurality of ribonucleoprotein enzymes.Annu Rev Biochem. 2002;71:165-89. doi: 10.1146/annurev.biochem.71.110601.135352. Epub 2001 Nov 9. Annu Rev Biochem. 2002. PMID: 12045094 Free PMC article. Review.
-
Functional characterization of the conserved amino acids in Pop1p, the largest common protein subunit of yeast RNases P and MRP.RNA. 2006 Jun;12(6):1023-37. doi: 10.1261/rna.23206. Epub 2006 Apr 17. RNA. 2006. PMID: 16618965 Free PMC article.
-
Footprinting analysis demonstrates extensive similarity between eukaryotic RNase P and RNase MRP holoenzymes.RNA. 2008 Aug;14(8):1558-67. doi: 10.1261/rna.1106408. Epub 2008 Jun 25. RNA. 2008. PMID: 18579867 Free PMC article.
-
Archaeal/eukaryal RNase P: subunits, functions and RNA diversification.Nucleic Acids Res. 2010 Dec;38(22):7885-94. doi: 10.1093/nar/gkq701. Epub 2010 Aug 16. Nucleic Acids Res. 2010. PMID: 20716516 Free PMC article. Review.
-
Structural and functional similarities between MRP and RNase P.Mol Biol Rep. 1995-1996;22(2-3):81-5. doi: 10.1007/BF00988710. Mol Biol Rep. 1995. PMID: 8901492 Review.
Cited by
-
The role of structure in regulatory RNA elements.Biosci Rep. 2024 Oct 30;44(10):BSR20240139. doi: 10.1042/BSR20240139. Biosci Rep. 2024. PMID: 39364891 Free PMC article. Review.
-
Identification of RMP24 and RMP64, human ribonuclease MRP-specific protein components.Cell Rep. 2025 Jun 24;44(6):115752. doi: 10.1016/j.celrep.2025.115752. Epub 2025 May 24. Cell Rep. 2025. PMID: 40413743 Free PMC article.
-
Discovery, structure, mechanisms, and evolution of protein-only RNase P enzymes.J Biol Chem. 2024 Mar;300(3):105731. doi: 10.1016/j.jbc.2024.105731. Epub 2024 Feb 8. J Biol Chem. 2024. PMID: 38336295 Free PMC article. Review.
-
Molecular determinants of RNase MRP specificity and function.bioRxiv [Preprint]. 2025 Jan 28:2025.01.28.635360. doi: 10.1101/2025.01.28.635360. bioRxiv. 2025. PMID: 39974906 Free PMC article. Preprint.
-
Structural basis of circularly permuted group II intron self-splicing.Nat Struct Mol Biol. 2025 Jun;32(6):1091-1100. doi: 10.1038/s41594-025-01484-x. Epub 2025 Jan 31. Nat Struct Mol Biol. 2025. PMID: 39890981
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
