

Amyloid-β aggregates activate peripheral monocytes in mild cognitive impairment
- PMID: 38336934
- PMCID: PMC10858199
- DOI: 10.1038/s41467-024-45627-y
Amyloid-β aggregates activate peripheral monocytes in mild cognitive impairment
Abstract
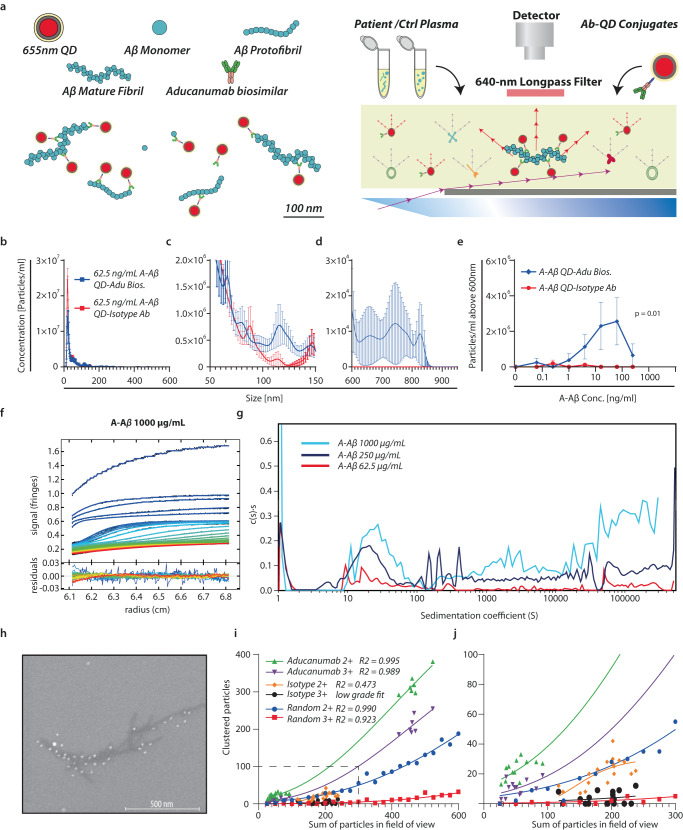
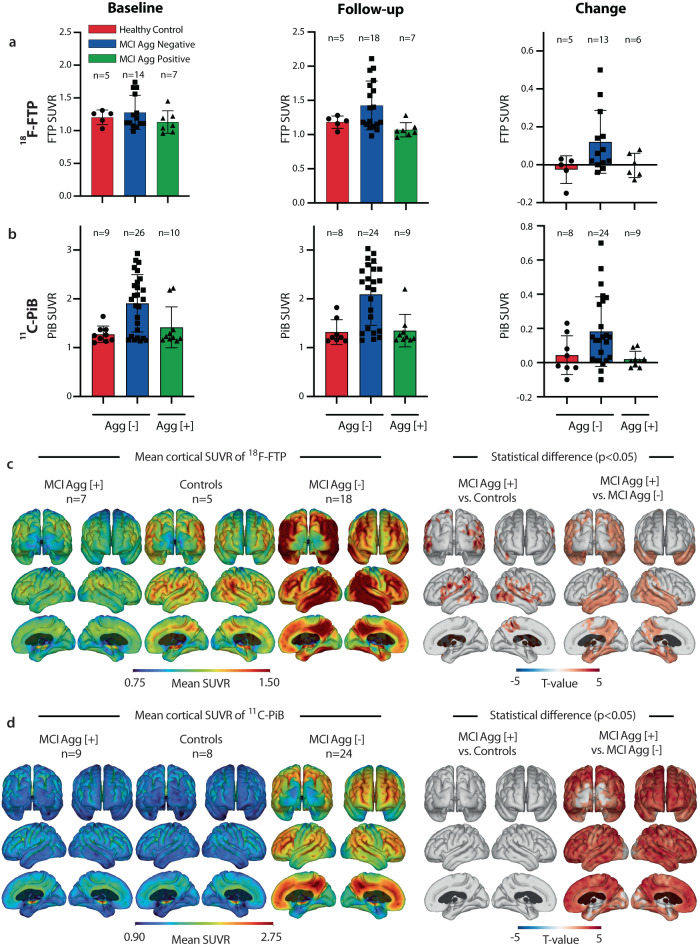
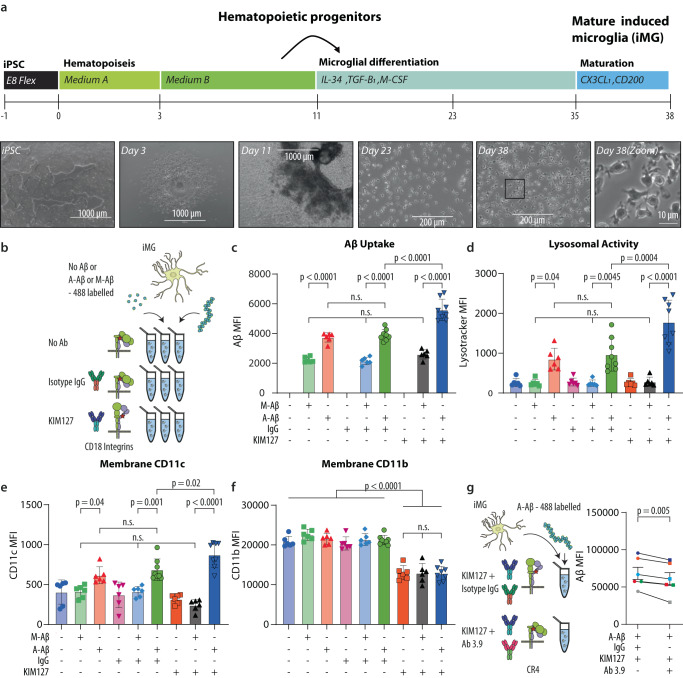
The peripheral immune system is important in neurodegenerative diseases, both in protecting and inflaming the brain, but the underlying mechanisms remain elusive. Alzheimer's Disease is commonly preceded by a prodromal period. Here, we report the presence of large Aβ aggregates in plasma from patients with mild cognitive impairment (n = 38). The aggregates are associated with low level Alzheimer's Disease-like brain pathology as observed by 11C-PiB PET and 18F-FTP PET and lowered CD18-rich monocytes. We characterize complement receptor 4 as a strong binder of amyloids and show Aβ aggregates are preferentially phagocytosed and stimulate lysosomal activity through this receptor in stem cell-derived microglia. KIM127 integrin activation in monocytes promotes size selective phagocytosis of Aβ. Hydrodynamic calculations suggest Aβ aggregates associate with vessel walls of the cortical capillaries. In turn, we hypothesize aggregates may provide an adhesion substrate for recruiting CD18-rich monocytes into the cortex. Our results support a role for complement receptor 4 in regulating amyloid homeostasis.
© 2024. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures





References
MeSH terms
Substances
Grants and funding
- AUFF-E-2015-FLS-9-6/Aarhus Universitets Forskningsfond (Aarhus University Research Foundation)
- AU IDEAS 2013/Aarhus Universitets Forskningsfond (Aarhus University Research Foundation)
- AUFF-E-2015-FLS-9-6/Aarhus Universitets Forskningsfond (Aarhus University Research Foundation)
- AU IDEAS 2013/Aarhus Universitets Forskningsfond (Aarhus University Research Foundation)
- AU IDEAS 2013/Aarhus Universitets Forskningsfond (Aarhus University Research Foundation)
- NNF19OC0058516/Novo Nordisk Fonden (Novo Nordisk Foundation)
- NNF19OC0058516/Novo Nordisk Fonden (Novo Nordisk Foundation)
- NNF21OC0071574/Novo Nordisk Fonden (Novo Nordisk Foundation)
- R380-2021-1326/Lundbeckfonden (Lundbeck Foundation)
- R380-2021-1326/Lundbeckfonden (Lundbeck Foundation)
- 2030-00002B/Det Frie Forskningsråd (Danish Council for Independent Research)
- 3101-00104B/Det Frie Forskningsråd (Danish Council for Independent Research)
- 2030-00002B/Det Frie Forskningsråd (Danish Council for Independent Research)
- 4004-00305/Det Frie Forskningsråd (Danish Council for Independent Research)
- 820636/EC | EU Framework Programme for Research and Innovation H2020 | H2020 European Institute of Innovation and Technology (H2020 The European Institute of Innovation and Technology)
- DNRF135/Danmarks Grundforskningsfond (Danish National Research Foundation)
LinkOut - more resources
Full Text Sources
Medical
