

Visualization of translation reorganization upon persistent ribosome collision stress in mammalian cells
- PMID: 38340715
- PMCID: PMC7615912
- DOI: 10.1016/j.molcel.2024.01.015
Visualization of translation reorganization upon persistent ribosome collision stress in mammalian cells
Abstract
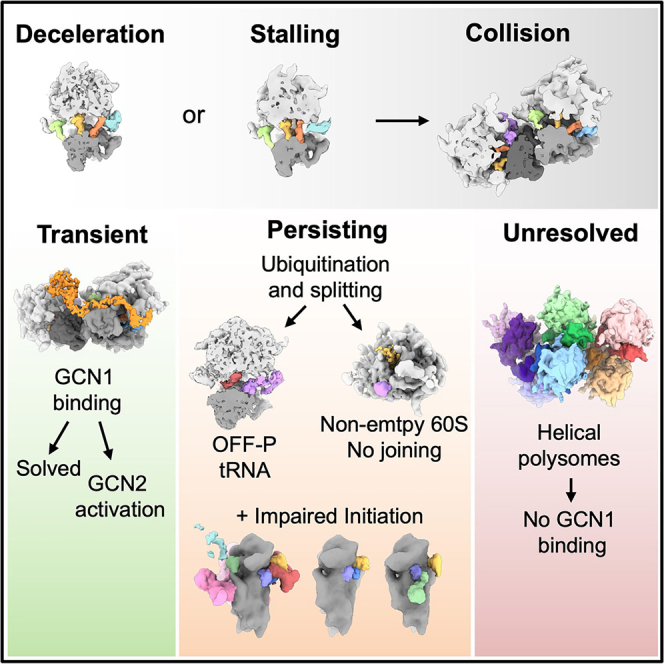
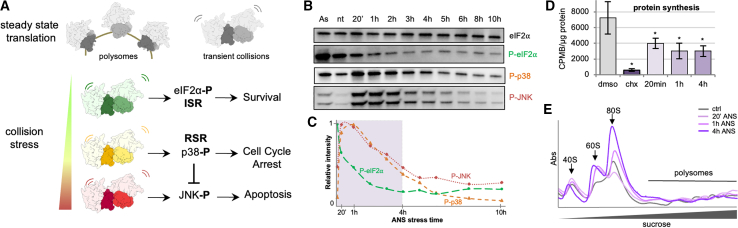
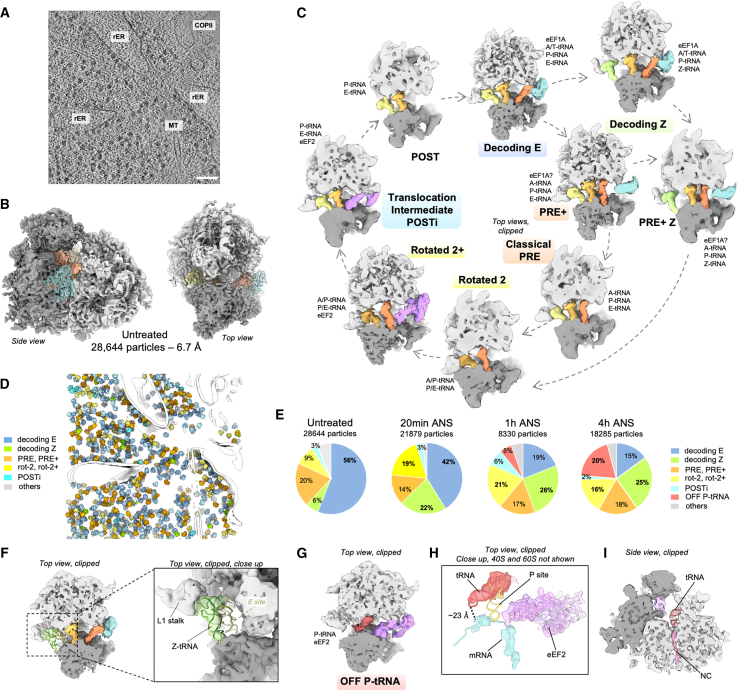
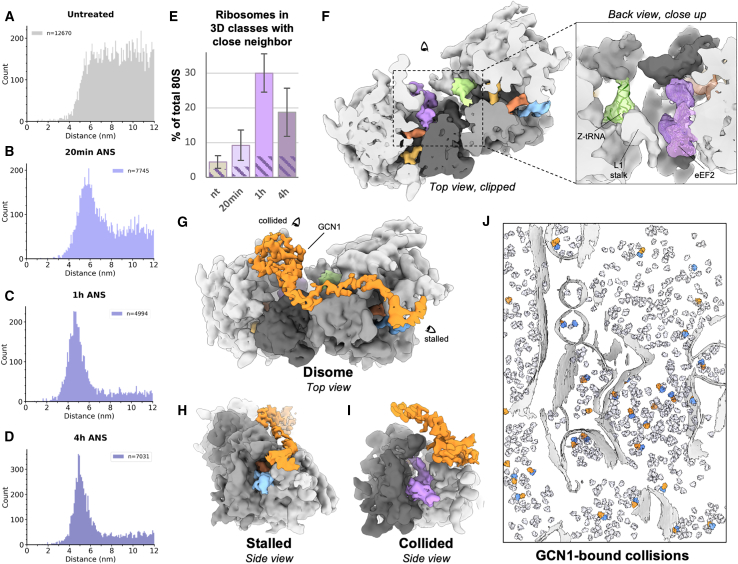
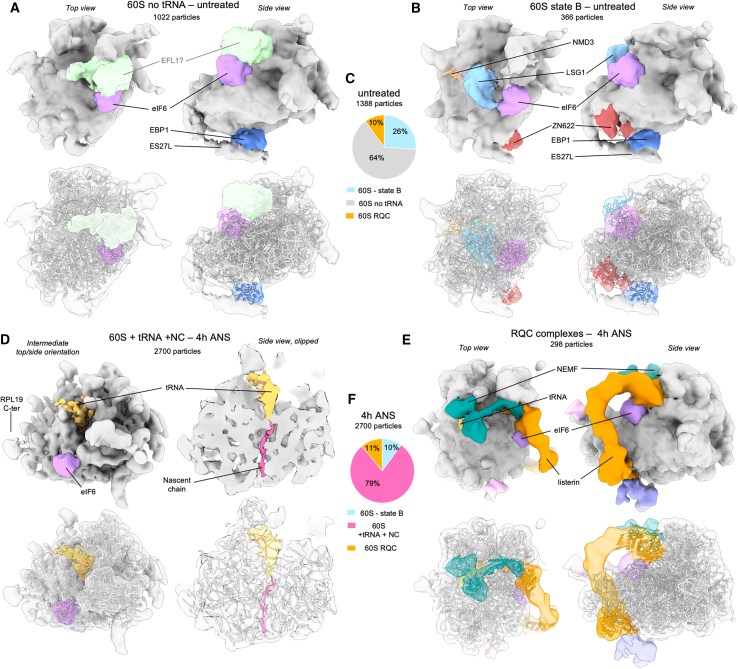
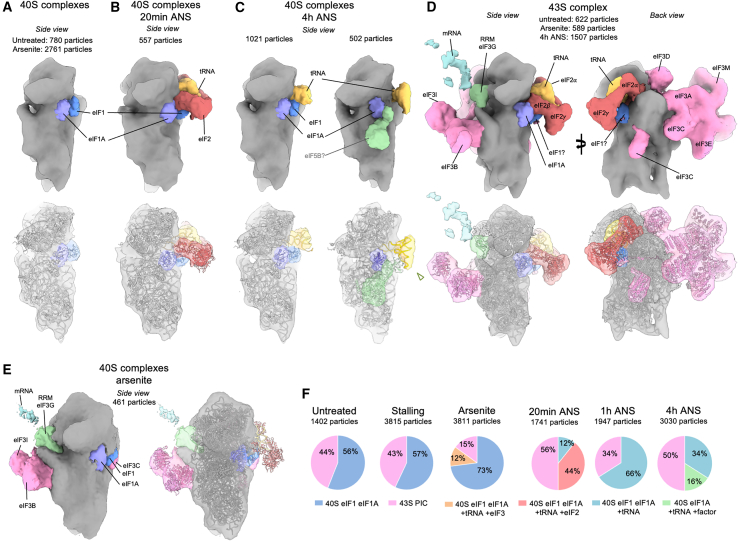
Aberrantly slow ribosomes incur collisions, a sentinel of stress that triggers quality control, signaling, and translation attenuation. Although each collision response has been studied in isolation, the net consequences of their collective actions in reshaping translation in cells is poorly understood. Here, we apply cryoelectron tomography to visualize the translation machinery in mammalian cells during persistent collision stress. We find that polysomes are compressed, with up to 30% of ribosomes in helical polysomes or collided disomes, some of which are bound to the stress effector GCN1. The native collision interface extends beyond the in vitro-characterized 40S and includes the L1 stalk and eEF2, possibly contributing to translocation inhibition. The accumulation of unresolved tRNA-bound 80S and 60S and aberrant 40S configurations identifies potentially limiting steps in collision responses. Our work provides a global view of the translation machinery in response to persistent collisions and a framework for quantitative analysis of translation dynamics in situ.
Keywords: cryoelectron tomography; initiation; polysome; ribosome collision; ribosome quality control; translation regulation.
Copyright © 2024 MRC Laboratory of Molecular Biology. Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests The authors declare no competing interests.
Figures
Update of
-
Visualization of translation reorganization upon persistent collision stress in mammalian cells.bioRxiv [Preprint]. 2023 Mar 24:2023.03.23.533914. doi: 10.1101/2023.03.23.533914. bioRxiv. 2023. Update in: Mol Cell. 2024 Mar 21;84(6):1078-1089.e4. doi: 10.1016/j.molcel.2024.01.015. PMID: 36993420 Free PMC article. Updated. Preprint.
References
-
- Narita M., Denk T., Matsuo Y., Sugiyama T., Kikuguchi C., Ito S., Sato N., Suzuki T., Hashimoto S., Machová I., et al. A distinct mammalian disome collision interface harbors K63-linked polyubiquitination of uS10 to trigger hRQT-mediated subunit dissociation. Nat. Commun. 2022;13:6411. - PMC - PubMed
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
