

A Dual-domain Engineered Antibody for Efficient HBV Suppression and Immune Responses Restoration
- PMID: 38342604
- PMCID: PMC11022716
- DOI: 10.1002/advs.202305316
A Dual-domain Engineered Antibody for Efficient HBV Suppression and Immune Responses Restoration
Abstract
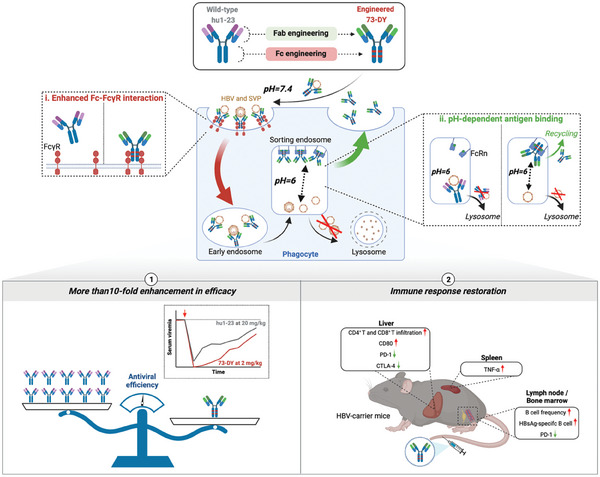
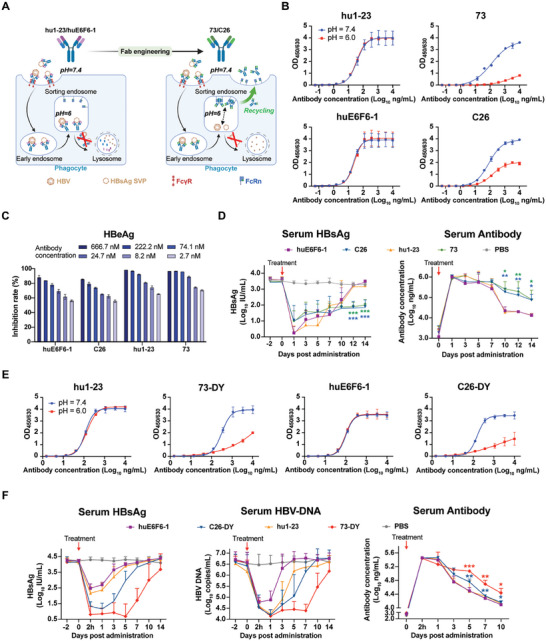
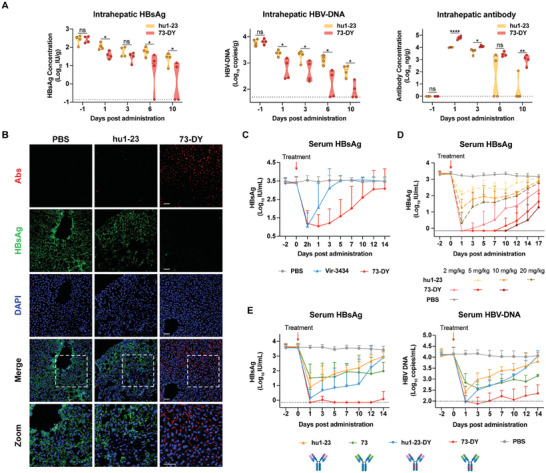
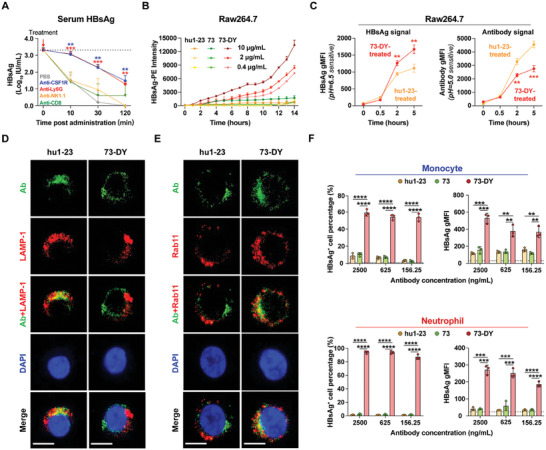
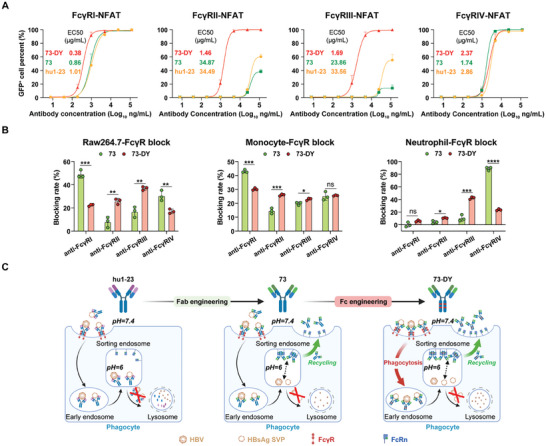
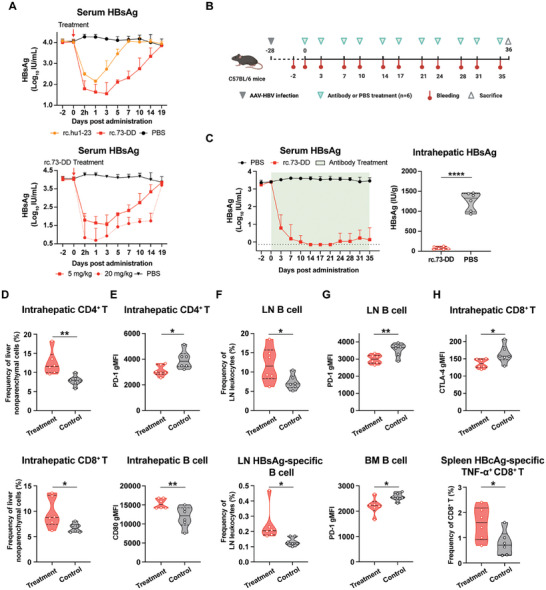
Chronic hepatitis B (CHB) remains a major public health concern because of the inefficiency of currently approved therapies in clearing the hepatitis B surface antigen (HBsAg). Antibody-based regimens have demonstrated potency regarding virus neutralization and HBsAg clearance. However, high dosages or frequent dosing are required for virologic control. In this study, a dual-domain-engineered anti-hepatitis B virus (HBV) therapeutic antibody 73-DY is developed that exhibits significantly improved efficacy regarding both serum and intrahepatic viral clearance. In HBV-tolerant mice, administration of a single dose of 73-DY at 2 mg kg-1 is sufficient to reduce serum HBsAg by over 3 log10 IU mL-1 and suppress HBsAg to < 100 IU mL-1 for two weeks, demonstrating a dose-lowering advantage of at least tenfold. Furthermore, 10 mg kg-1 of 73-DY sustainably suppressed serum viral levels to undetectable levels for ≈ 2 weeks. Molecular analyses indicate that the improved efficacy exhibited by 73-DY is attributable to the synergy between fragment antigen binding (Fab) and fragment crystallizable (Fc) engineering, which conferred sustained viral suppression and robust viral eradication, respectively. Long-term immunotherapy with reverse chimeric 73-DY facilitated the restoration of anti-HBV immune responses. This study provides a foundation for the development of next-generation antibody-based CHB therapies.
Keywords: antibody‐based immunotherapy; chronic hepatitis B; immune restoration; therapeutic efficacy.
© 2024 The Authors. Advanced Science published by Wiley‐VCH GmbH.
Conflict of interest statement
The author declares no conflicts of interest.
Figures
Similar articles
-
Antibody-mediated immunotherapy against chronic hepatitis B virus infection.Hum Vaccin Immunother. 2017 Aug 3;13(8):1768-1773. doi: 10.1080/21645515.2017.1319021. Epub 2017 May 19. Hum Vaccin Immunother. 2017. PMID: 28521640 Free PMC article.
-
Prolonged suppression of HBV in mice by a novel antibody that targets a unique epitope on hepatitis B surface antigen.Gut. 2016 Apr;65(4):658-71. doi: 10.1136/gutjnl-2014-308964. Epub 2015 Sep 30. Gut. 2016. PMID: 26423112
-
A Single Dose of Anti-HBsAg Antibody-Encoding mRNA-LNPs Suppressed HBsAg Expression: a Potential Cure of Chronic Hepatitis B Virus Infection.mBio. 2022 Aug 30;13(4):e0161222. doi: 10.1128/mbio.01612-22. Epub 2022 Jul 7. mBio. 2022. PMID: 35862767 Free PMC article.
-
Restoration of a functional antiviral immune response to chronic HBV infection by reducing viral antigen load: if not sufficient, is it necessary?Emerg Microbes Infect. 2021 Dec;10(1):1545-1554. doi: 10.1080/22221751.2021.1952851. Emerg Microbes Infect. 2021. PMID: 34227927 Free PMC article. Review.
-
A role for immune modulation in achieving functional cure for chronic hepatitis B among current changes in the landscape of new treatments.Expert Rev Gastroenterol Hepatol. 2023 Jul-Dec;17(11):1135-1147. doi: 10.1080/17474124.2023.2268503. Epub 2023 Nov 23. Expert Rev Gastroenterol Hepatol. 2023. PMID: 37847193 Review.
Cited by
-
Protein-Based Degraders: From Chemical Biology Tools to Neo-Therapeutics.Chem Rev. 2025 Feb 26;125(4):2120-2183. doi: 10.1021/acs.chemrev.4c00595. Epub 2025 Jan 17. Chem Rev. 2025. PMID: 39818743 Review.
-
HBsAg and TLR7/8 dual-targeting antibody-drug conjugates induce sustained anti-HBV activity in AAV/HBV mice: a preliminary study.Antib Ther. 2024 Jul 3;7(3):249-255. doi: 10.1093/abt/tbae016. eCollection 2024 Jul. Antib Ther. 2024. PMID: 39262443 Free PMC article.
References
-
- Asrani S., Devarbhavi H., Eaton J., Kamath P., J Hepatol 2019, 70, 151. - PubMed
-
- Tang L., Covert E., Wilson E., Kottilil S., JAMA, J. Am. Med. Assoc. 2018, 319, 1802. - PubMed
-
- World Health Organization . Hepatitis B. 2023 July 18 [cited 2023 October]; Available from: https://www.who.int/en/news‐room/fact‐sheets/detail/hepatitis‐b.
-
- Jeng W., Papatheodoridis G., Lok A., Lancet 2023, 401, 1039. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources