

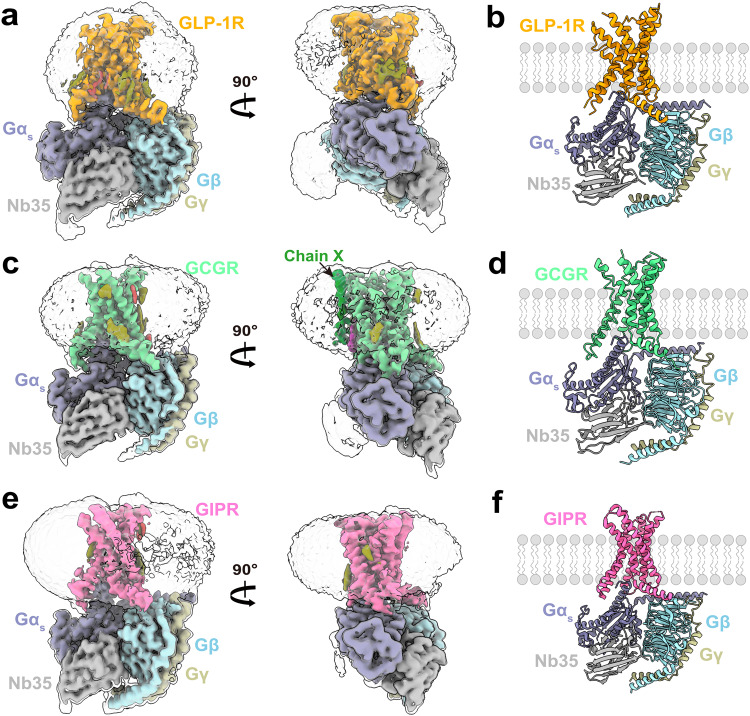
Molecular features of the ligand-free GLP-1R, GCGR and GIPR in complex with Gs proteins
- PMID: 38346960
- PMCID: PMC10861504
- DOI: 10.1038/s41421-024-00649-0
Molecular features of the ligand-free GLP-1R, GCGR and GIPR in complex with Gs proteins
Abstract
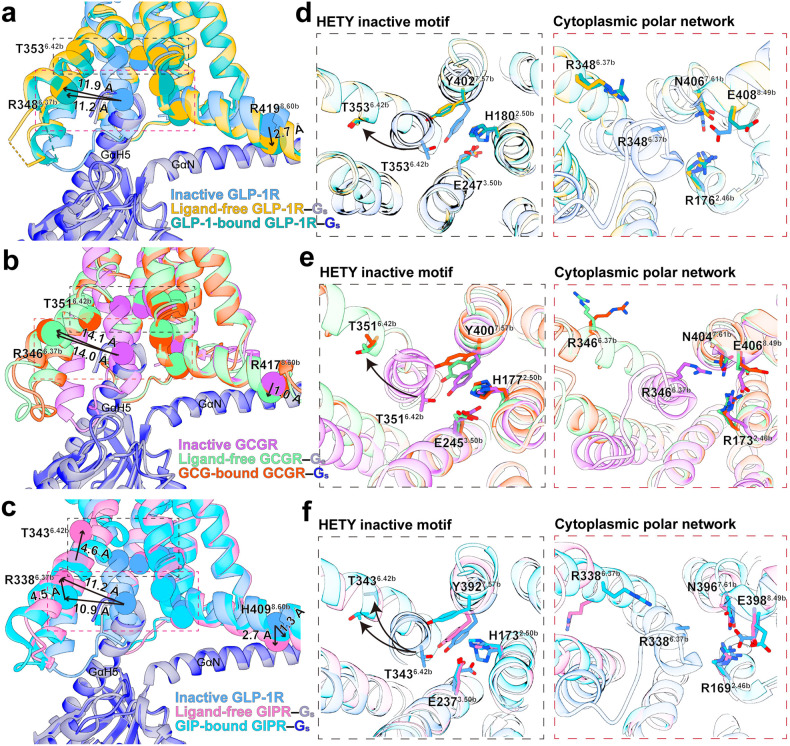
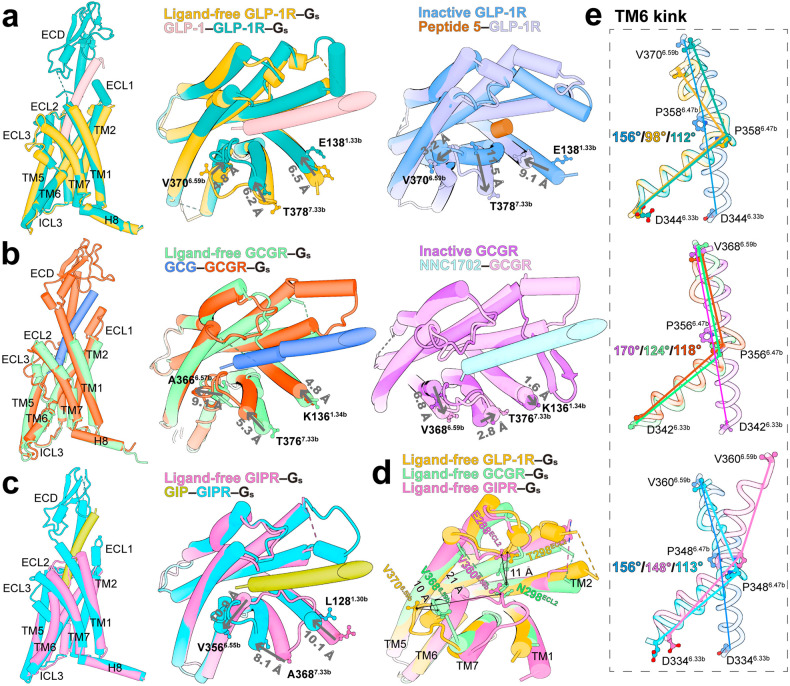
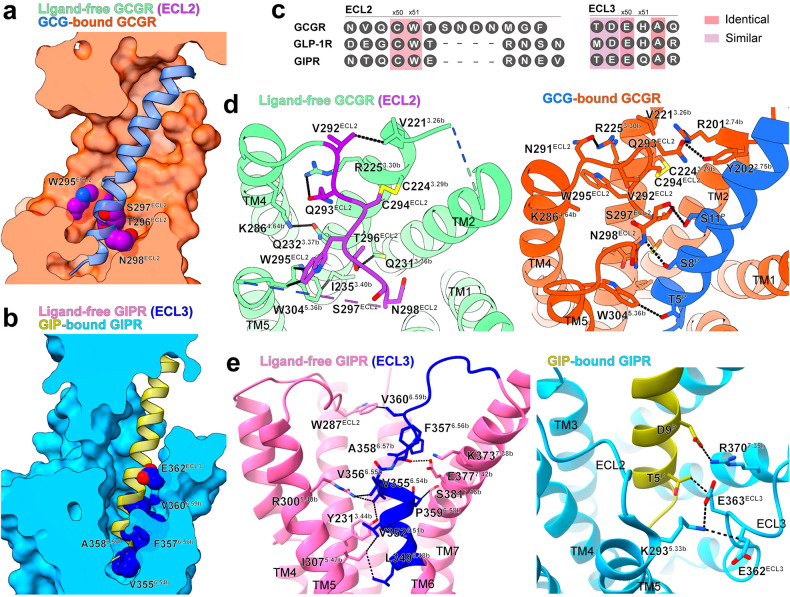
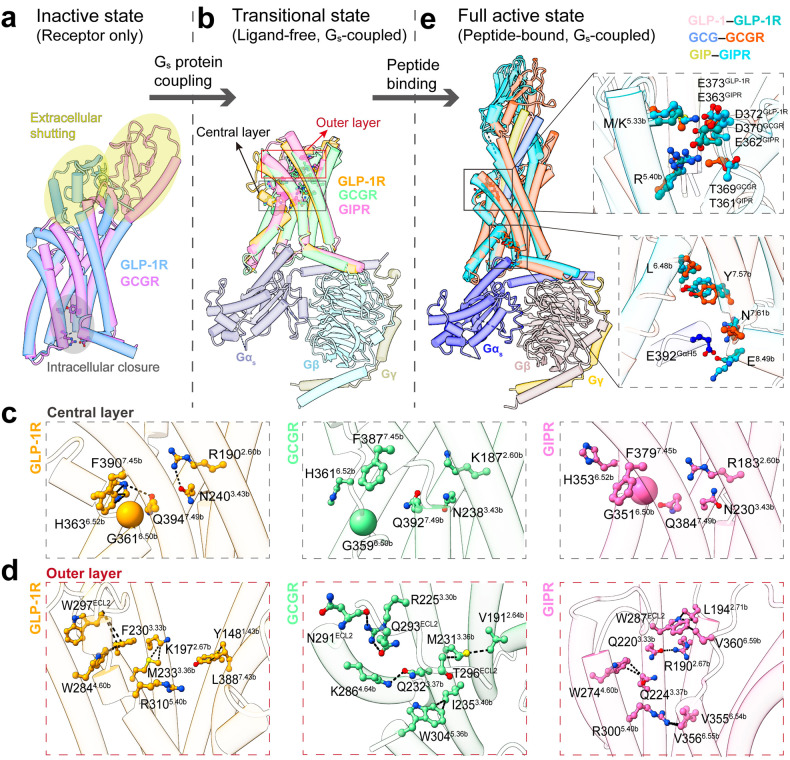
Class B1 G protein-coupled receptors (GPCRs) are important regulators of many physiological functions such as glucose homeostasis, which is mainly mediated by three peptide hormones, i.e., glucagon-like peptide-1 (GLP-1), glucagon (GCG), and glucose-dependent insulinotropic polypeptide (GIP). They trigger a cascade of signaling events leading to the formation of an active agonist-receptor-G protein complex. However, intracellular signal transducers can also activate the receptor independent of extracellular stimuli, suggesting an intrinsic role of G proteins in this process. Here, we report cryo-electron microscopy structures of the human GLP-1 receptor (GLP-1R), GCG receptor (GCGR), and GIP receptor (GIPR) in complex with Gs proteins without the presence of cognate ligands. These ligand-free complexes share a similar intracellular architecture to those bound by endogenous peptides, in which, the Gs protein alone directly opens the intracellular binding cavity and rewires the extracellular orthosteric pocket to stabilize the receptor in a state unseen before. While the peptide-binding site is partially occupied by the inward folded transmembrane helix 6 (TM6)-extracellular loop 3 (ECL3) juncture of GIPR or a segment of GCGR ECL2, the extracellular portion of GLP-1R adopts a conformation close to the active state. Our findings offer valuable insights into the distinct activation mechanisms of these three important receptors. It is possible that in the absence of a ligand, the intracellular half of transmembrane domain is mobilized with the help of Gs protein, which in turn rearranges the extracellular half to form a transitional conformation, facilitating the entry of the peptide N-terminus.
© 2024. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures
Similar articles
-
Human epicardial adipose tissue expresses glucose-dependent insulinotropic polypeptide, glucagon, and glucagon-like peptide-1 receptors as potential targets of pleiotropic therapies.Eur J Prev Cardiol. 2023 Jun 1;30(8):680-693. doi: 10.1093/eurjpc/zwad050. Eur J Prev Cardiol. 2023. PMID: 36799940
-
Cryo-electron microscopy structure of the glucagon receptor with a dual-agonist peptide.J Biol Chem. 2020 Jul 10;295(28):9313-9325. doi: 10.1074/jbc.RA120.013793. Epub 2020 May 5. J Biol Chem. 2020. PMID: 32371397 Free PMC article.
-
Structural analysis of the dual agonism at GLP-1R and GCGR.Proc Natl Acad Sci U S A. 2023 Aug 15;120(33):e2303696120. doi: 10.1073/pnas.2303696120. Epub 2023 Aug 7. Proc Natl Acad Sci U S A. 2023. PMID: 37549266 Free PMC article.
-
GLP-1 and GIP receptor signaling in beta cells - A review of receptor interactions and co-stimulation.Peptides. 2022 May;151:170749. doi: 10.1016/j.peptides.2022.170749. Epub 2022 Jan 19. Peptides. 2022. PMID: 35065096 Review.
-
Insights into the structure and activation mechanism of some class B1 GPCR family members.Mol Biol Rep. 2024 Sep 6;51(1):966. doi: 10.1007/s11033-024-09876-w. Mol Biol Rep. 2024. PMID: 39240462 Review.
Cited by
-
Exploring Conformational Transitions in Biased and Balanced Ligand Binding of GLP-1R.Molecules. 2025 Jul 31;30(15):3216. doi: 10.3390/molecules30153216. Molecules. 2025. PMID: 40807391 Free PMC article.
-
Structural insights into GPCR signaling activated by peptide ligands: from molecular mechanism to therapeutic application.Exp Mol Med. 2025 Jul;57(7):1467-1481. doi: 10.1038/s12276-025-01497-y. Epub 2025 Jul 8. Exp Mol Med. 2025. PMID: 40629042 Free PMC article. Review.
-
CXC Chemokine Ligand 12 Facilitates Gi Protein Binding to CXC Chemokine Receptor 4 by Stabilizing Packing of the Proline-Isoleucine-Phenylalanine Motif: Insights from Automated Path Searching.J Am Chem Soc. 2025 Mar 26;147(12):10129-10138. doi: 10.1021/jacs.4c14293. Epub 2025 Mar 17. J Am Chem Soc. 2025. PMID: 40096846 Free PMC article.
-
The dual GLP-1 and GIP receptor agonist tirzapetide provides an unintended interaction with the β-adrenoceptors and plays a role in glucose metabolism in hyperglycemic or senescent cardiac cells.Cardiovasc Diabetol. 2025 Aug 18;24(1):338. doi: 10.1186/s12933-025-02828-z. Cardiovasc Diabetol. 2025. PMID: 40826463 Free PMC article.
-
Oral FPR2/ALX modulators tune myeloid cell activity to ameliorate mucosal inflammation in inflammatory bowel disease.Acta Pharmacol Sin. 2025 Jul;46(7):1958-1973. doi: 10.1038/s41401-025-01525-7. Epub 2025 Mar 11. Acta Pharmacol Sin. 2025. PMID: 40069490
References
Grants and funding
- 81872915; 82073904; 82273961/National Natural Science Foundation of China (National Science Foundation of China)
- 32200576/National Natural Science Foundation of China (National Science Foundation of China)
- 82204474/National Natural Science Foundation of China (National Science Foundation of China)
- 82273985; 82121005; 81973373/National Natural Science Foundation of China (National Science Foundation of China)
- 2022M710806/China Postdoctoral Science Foundation
LinkOut - more resources
Full Text Sources
