

Species-specific roles of the Notch ligands, receptors, and targets orchestrating the signaling landscape of the segmentation clock
- PMID: 38348091
- PMCID: PMC10859470
- DOI: 10.3389/fcell.2023.1327227
Species-specific roles of the Notch ligands, receptors, and targets orchestrating the signaling landscape of the segmentation clock
Abstract
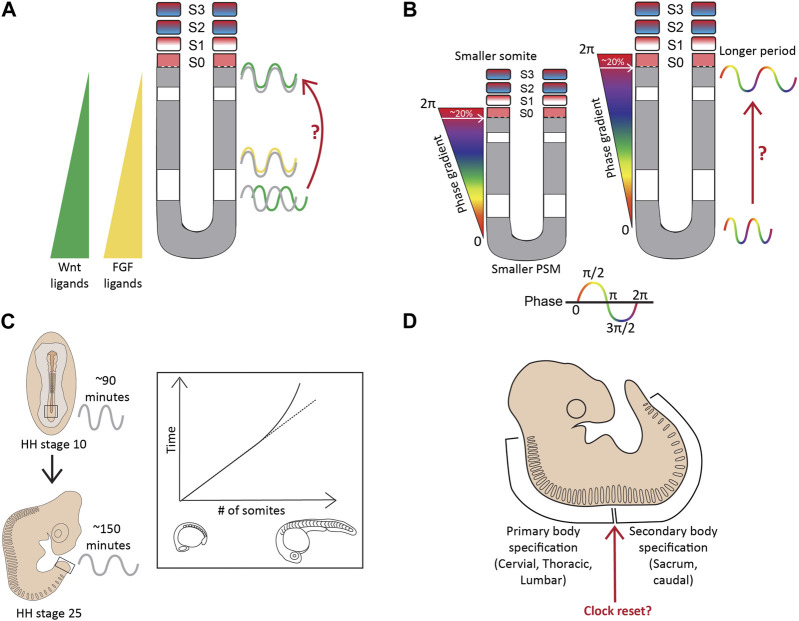
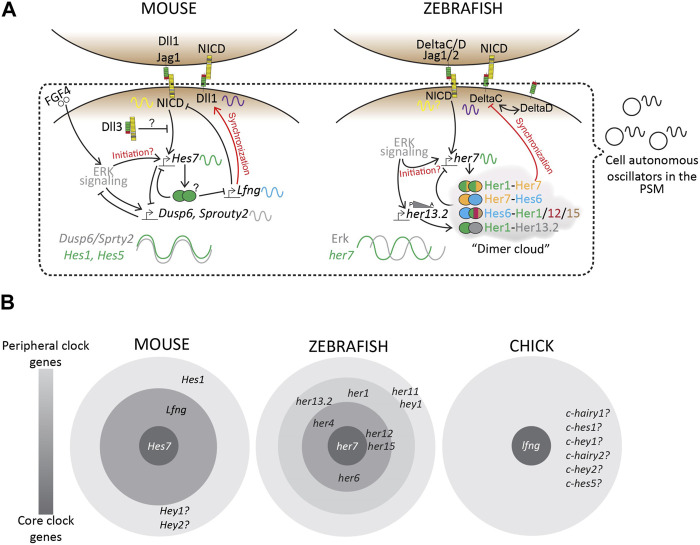
Somitogenesis is a hallmark feature of all vertebrates and some invertebrate species that involves the periodic formation of block-like structures called somites. Somites are transient embryonic segments that eventually establish the entire vertebral column. A highly conserved molecular oscillator called the segmentation clock underlies this periodic event and the pace of this clock regulates the pace of somite formation. Although conserved signaling pathways govern the clock in most vertebrates, the mechanisms underlying the species-specific divergence in various clock characteristics remain elusive. For example, the segmentation clock in classical model species such as zebrafish, chick, and mouse embryos tick with a periodicity of ∼30, ∼90, and ∼120 min respectively. This enables them to form the species-specific number of vertebrae during their overall timespan of somitogenesis. Here, we perform a systematic review of the species-specific features of the segmentation clock with a keen focus on mouse embryos. We perform this review using three different perspectives: Notch-responsive clock genes, ligand-receptor dynamics, and synchronization between neighboring oscillators. We further review reports that use non-classical model organisms and in vitro model systems that complement our current understanding of the segmentation clock. Our review highlights the importance of comparative developmental biology to further our understanding of this essential developmental process.
Keywords: Notch signailing pathway; gene oscillation; presomitic mesoderm (PSM); segmentation clock; somite; somitogenesis.
Copyright © 2024 Ramesh and Chu.
Conflict of interest statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Figures





Similar articles
-
Oscillatory gene expression and somitogenesis.Wiley Interdiscip Rev Dev Biol. 2012 Sep-Oct;1(5):629-41. doi: 10.1002/wdev.46. Epub 2012 Mar 22. Wiley Interdiscip Rev Dev Biol. 2012. PMID: 23799565 Review.
-
The vertebrate segmentation clock.J Anat. 2001 Jul-Aug;199(Pt 1-2):169-75. doi: 10.1046/j.1469-7580.2001.19910169.x. J Anat. 2001. PMID: 11523819 Free PMC article. Review.
-
Notch signalling and the synchronization of the somite segmentation clock.Nature. 2000 Nov 23;408(6811):475-9. doi: 10.1038/35044091. Nature. 2000. PMID: 11100729
-
Comparative analysis of her genes during fish somitogenesis suggests a mouse/chick-like mode of oscillation in medaka.Dev Genes Evol. 2006 Jun;216(6):315-32. doi: 10.1007/s00427-006-0059-6. Epub 2006 Mar 17. Dev Genes Evol. 2006. PMID: 16544152
-
Somite boundary determination in normal and clock-less vertebrate embryos.Dev Growth Differ. 2020 Apr;62(3):177-187. doi: 10.1111/dgd.12655. Epub 2020 Feb 28. Dev Growth Differ. 2020. PMID: 32108939 Review.
Cited by
-
NOTCH1 S2513 is critical for the regulation of NICD levels impacting the segmentation clock in hiPSC-derived PSM cells and somitoids.Genes Dev. 2025 Sep 2;39(17-18):1025-1044. doi: 10.1101/gad.352909.125. Genes Dev. 2025. PMID: 40441890 Free PMC article.
References
Publication types
LinkOut - more resources
Full Text Sources
Miscellaneous