

RIP-seq reveals RNAs that interact with RNA polymerase and primary sigma factors in bacteria
- PMID: 38348908
- PMCID: PMC11077062
- DOI: 10.1093/nar/gkae081
RIP-seq reveals RNAs that interact with RNA polymerase and primary sigma factors in bacteria
Abstract
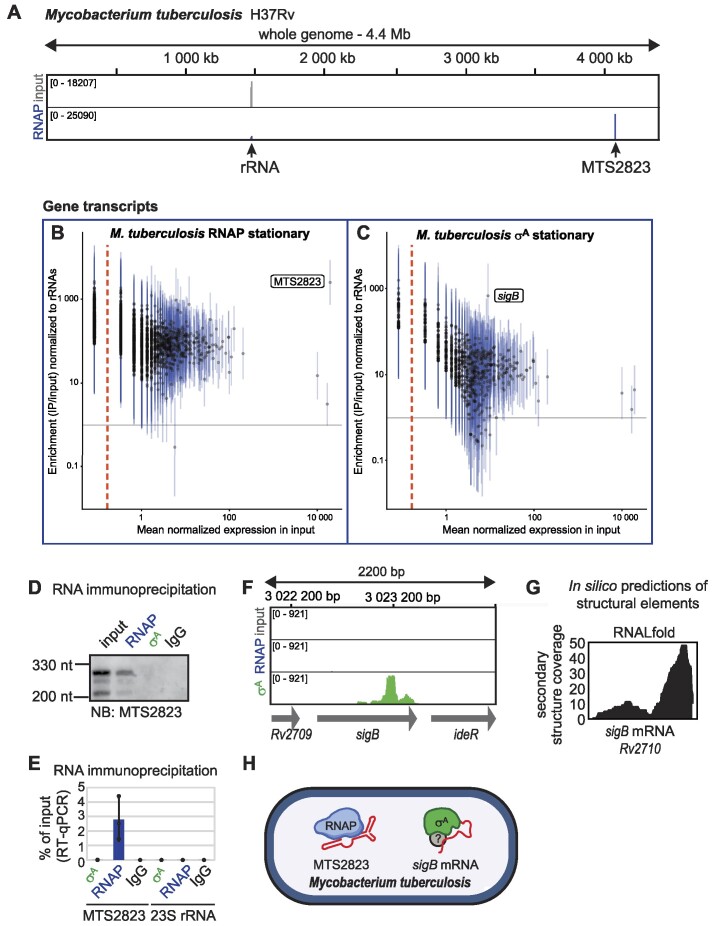
Bacteria have evolved structured RNAs that can associate with RNA polymerase (RNAP). Two of them have been known so far-6S RNA and Ms1 RNA but it is unclear if any other types of RNAs binding to RNAP exist in bacteria. To identify all RNAs interacting with RNAP and the primary σ factors, we have established and performed native RIP-seq in Bacillus subtilis, Corynebacterium glutamicum, Streptomyces coelicolor, Mycobacterium smegmatis and the pathogenic Mycobacterium tuberculosis. Besides known 6S RNAs in B. subtilis and Ms1 in M. smegmatis, we detected MTS2823, a homologue of Ms1, on RNAP in M. tuberculosis. In C. glutamicum, we discovered novel types of structured RNAs that associate with RNAP. Furthermore, we identified other species-specific RNAs including full-length mRNAs, revealing a previously unknown landscape of RNAs interacting with the bacterial transcription machinery.
© The Author(s) 2024. Published by Oxford University Press on behalf of Nucleic Acids Research.
Figures











References
-
- Sensi P. History of the development of rifampin. Rev. Infect. Dis. 1983; 5:S402–S406. - PubMed
-
- Gomez M., Doukhan L., Nair G., Smith I.. sigA is an essential gene in Mycobacterium smegmatis. Mol. Microbiol. 1998; 29:617–628. - PubMed
-
- Brown K.L., Wood S., Buttner M.J.. Isolation and characterization of the major vegetative RNA polymerase of Streptomyces coelicolor A3(2); renaturation of a sigma subunit using GroEL. Mol. Microbiol. 1992; 6:1133–1139. - PubMed
-
- Browning D.F., Busby S.J.. Local and global regulation of transcription initiation in bacteria. Nat. Rev. Micro. 2016; 14:638–650. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
