Selenium reduction of ubiquinone via SQOR suppresses ferroptosis
- PMID: 38351124
- PMCID: PMC11694790
- DOI: 10.1038/s42255-024-00974-4
Selenium reduction of ubiquinone via SQOR suppresses ferroptosis
Abstract
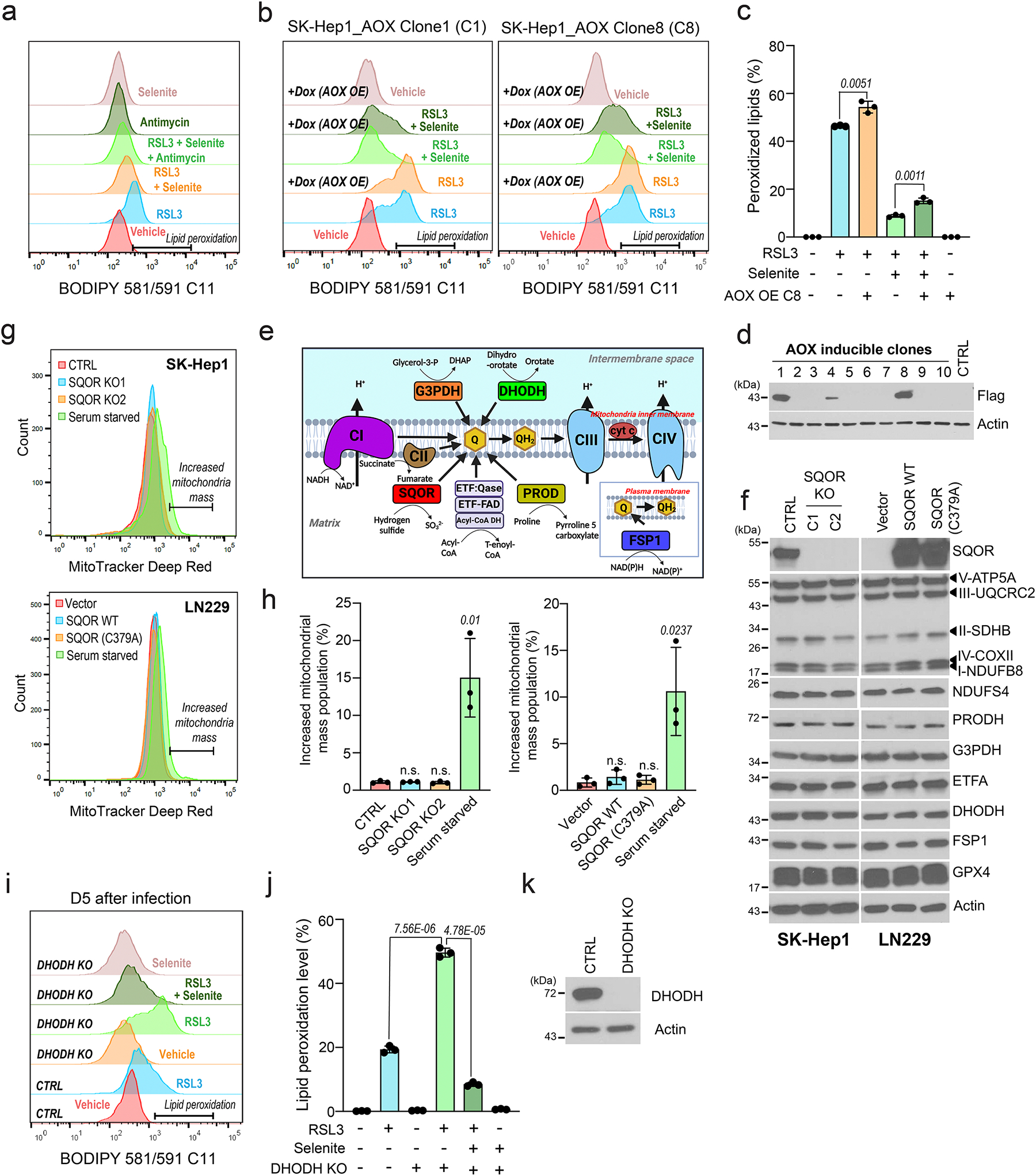
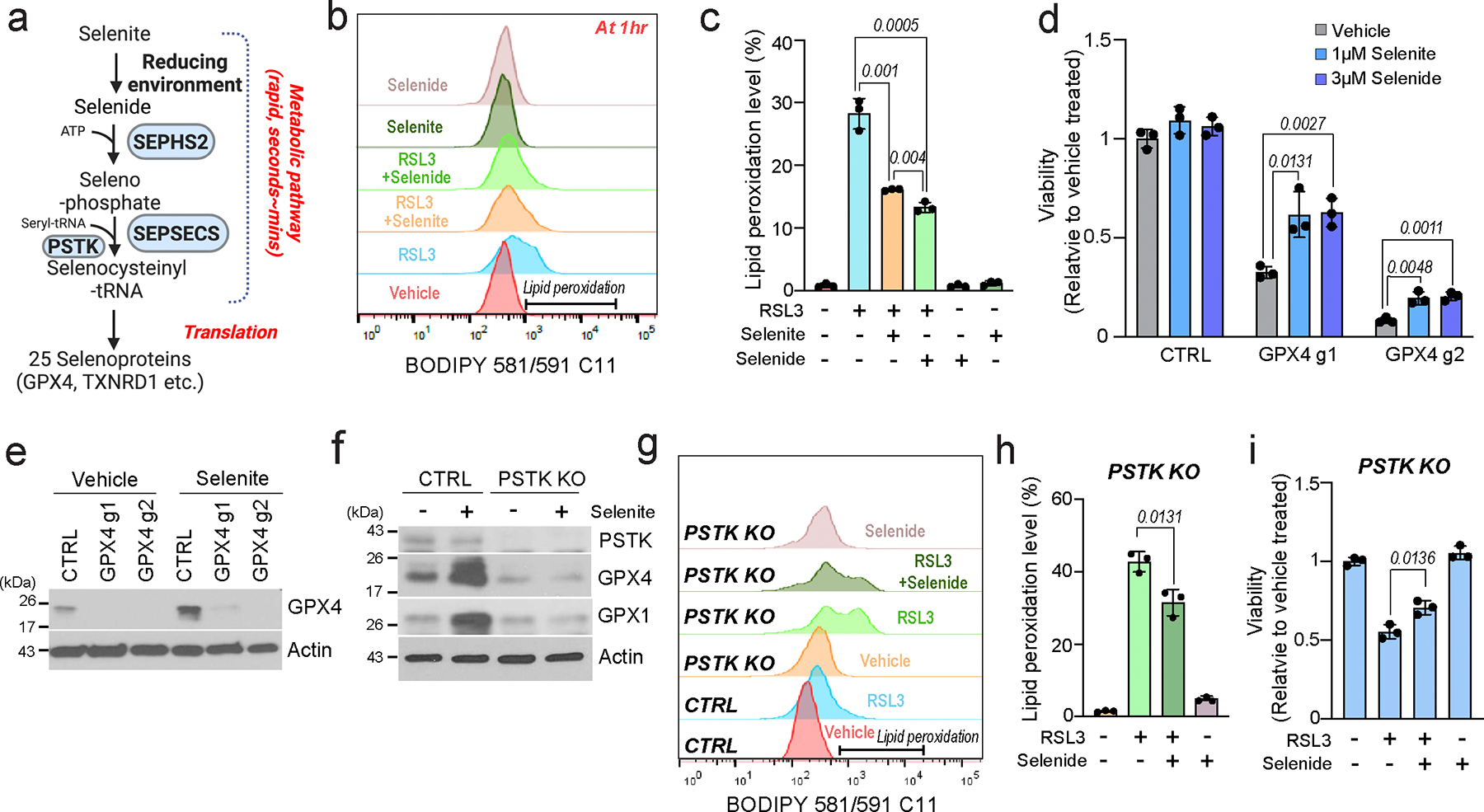
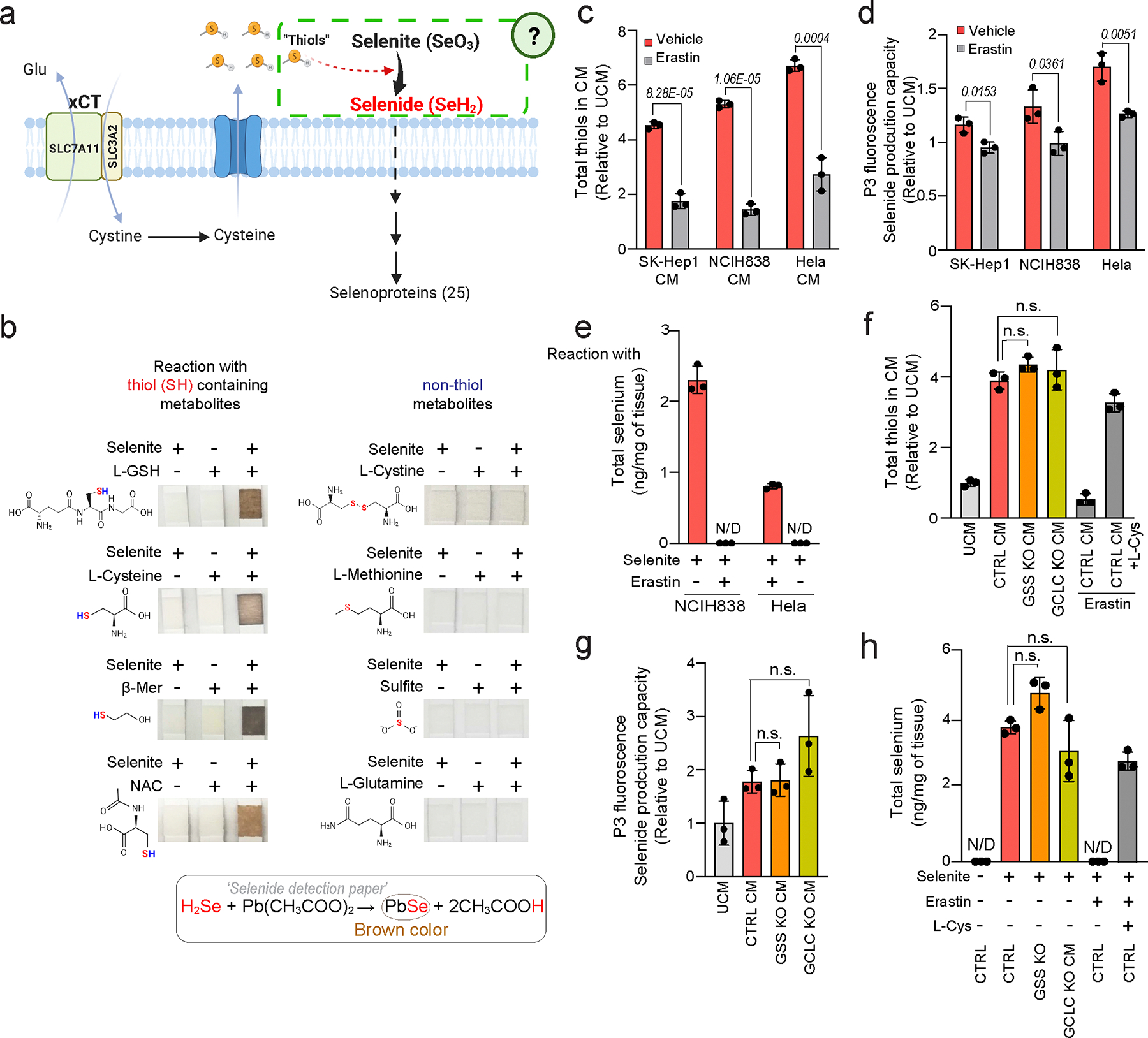
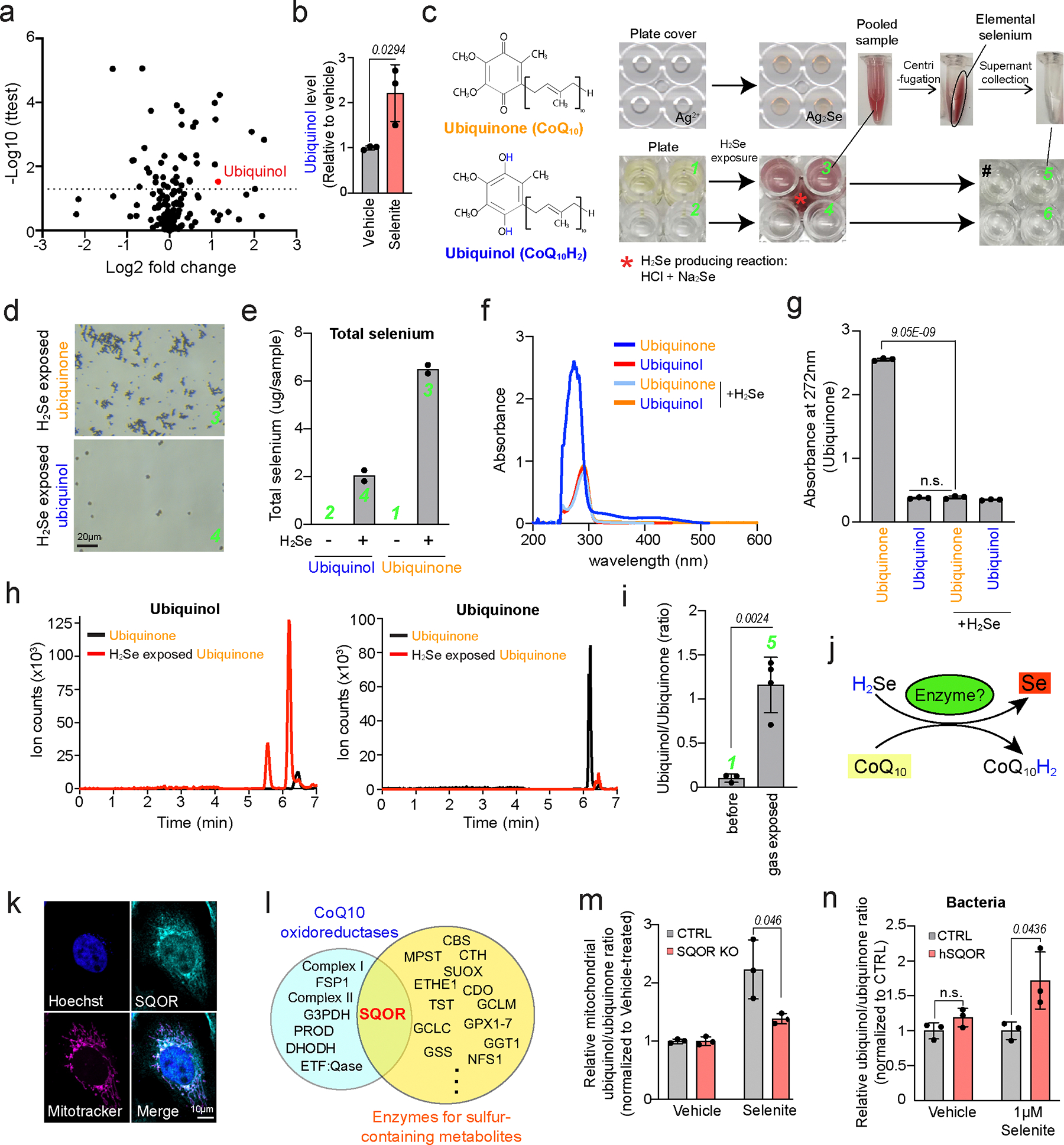
The canonical biological function of selenium is in the production of selenocysteine residues of selenoproteins, and this forms the basis for its role as an essential antioxidant and cytoprotective micronutrient. Here we demonstrate that, via its metabolic intermediate hydrogen selenide, selenium reduces ubiquinone in the mitochondria through catalysis by sulfide quinone oxidoreductase. Through this mechanism, selenium rapidly protects against lipid peroxidation and ferroptosis in a timescale that precedes selenoprotein production, doing so even when selenoprotein production has been eliminated. Our findings identify a regulatory mechanism against ferroptosis that implicates sulfide quinone oxidoreductase and expands our understanding of selenium in biology.
© 2024. The Author(s), under exclusive licence to Springer Nature Limited.
Conflict of interest statement
Competing interests statement
J.A.O. is a member of the scientific advisory board for Vicinitas Therapeutics and has ferroptosis-related patent applications. The remaining authors declare no competing interests.
Figures