

Targeting yeast topoisomerase II by imidazo and triazoloacridinone derivatives resulting in their antifungal activity
- PMID: 38351313
- PMCID: PMC10864382
- DOI: 10.1038/s41598-024-54252-0
Targeting yeast topoisomerase II by imidazo and triazoloacridinone derivatives resulting in their antifungal activity
Abstract
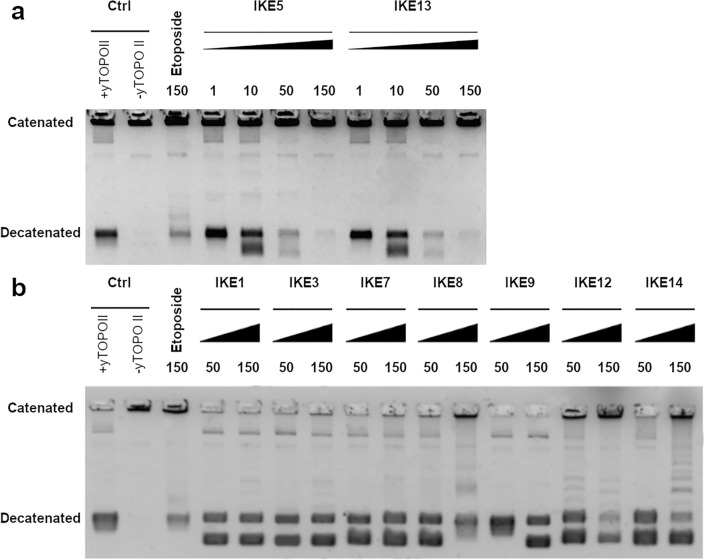
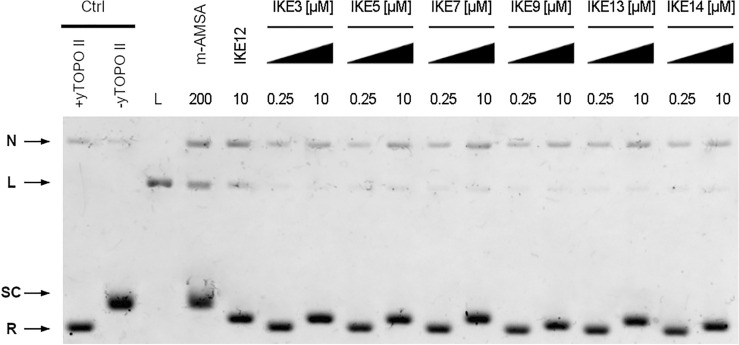
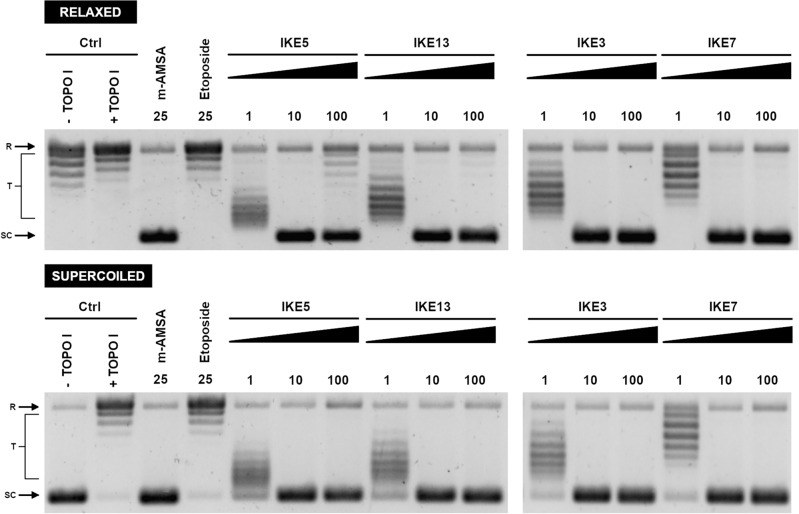
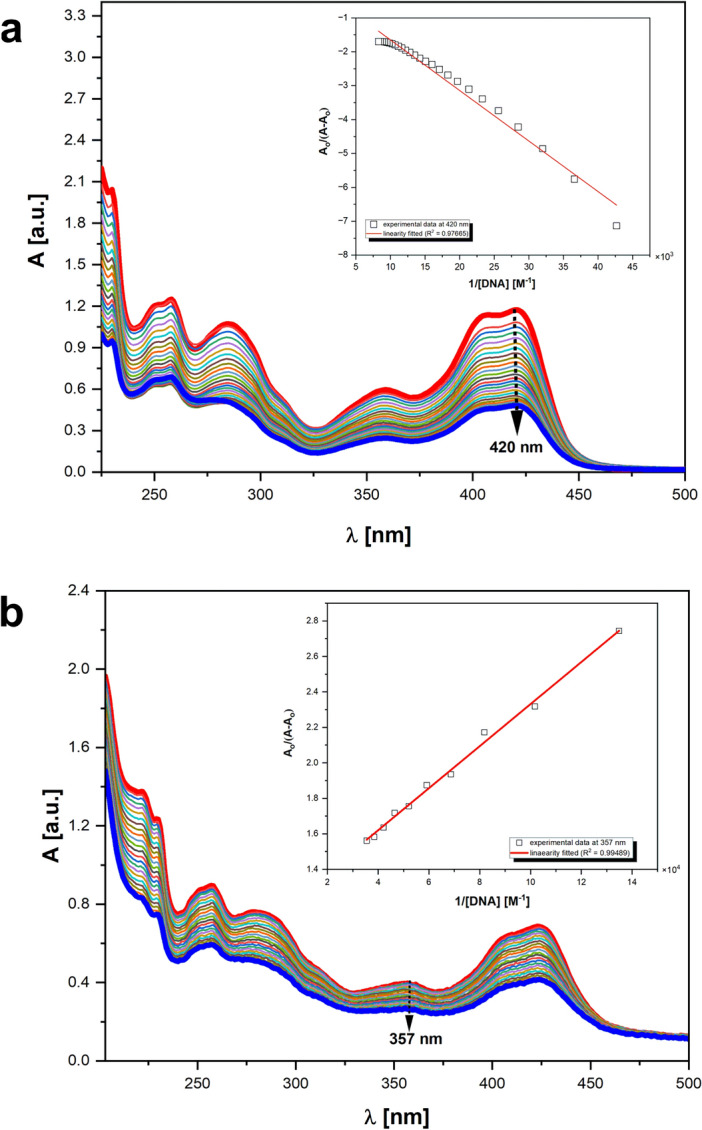
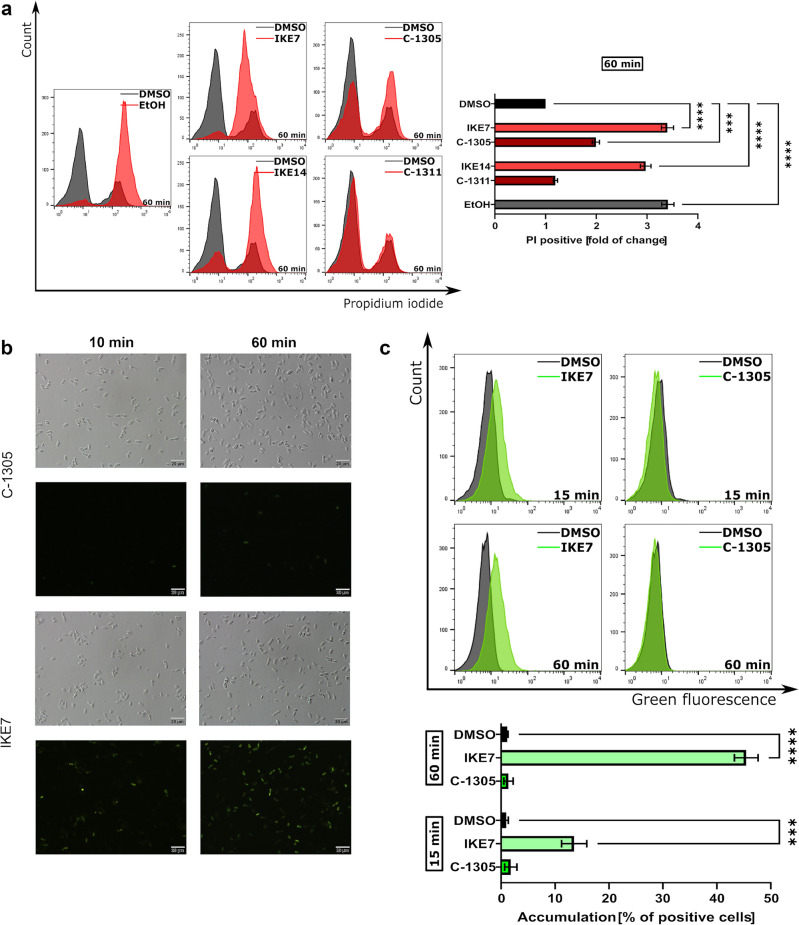
Fungal pathogens are considered as serious factors for deadly diseases and are a case of medical concern. Invasive fungal infections also complicate the clinical course of COVID-19, leading to a significant increase in mortality. Furthermore, fungal strains' multidrug resistance has increased the demand for antifungals with a different mechanism of action. The present study aimed to identify antifungal compounds targeting yeast topoisomerase II (yTOPOII) derived from well-known human topoisomerase II (hTOPOII) poisons C-1305 and C-1311. Two sets of derivatives: triazoloacridinones (IKE1-8) and imidazoacridinones (IKE9-14) were synthetized and evaluated with a specific emphasis on the molecular mechanism of action. Our results indicated that their effectiveness as enzyme inhibitors was not solely due to intercalation ability but also as a result of influence on catalytic activity by the formation of covalent complexes between plasmid DNA and yTOPOII. Lysine conjunction increased the strength of the compound's interaction with DNA and improved penetration into the fungal cells. Triazoloacridinone derivatives in contrast to starting compound C-1305 exhibited moderate antifungal activity and at least twice lower cytotoxicity. Importantly, compounds (IKE5-8) were not substrates for multidrug ABC transporters whereas a derivative conjugated with lysine (IKE7), showed the ability to overcome C. glabrata fluconazole-resistance (MIC 32-64 µg mL-1).
© 2024. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures
Similar articles
-
Antifungal Activity of Fibrate-Based Compounds and Substituted Pyrroles That Inhibit the Enzyme 3-Hydroxy-methyl-glutaryl-CoA Reductase of Candida glabrata (CgHMGR), Thus Decreasing Yeast Viability and Ergosterol Synthesis.Microbiol Spectr. 2022 Apr 27;10(2):e0164221. doi: 10.1128/spectrum.01642-21. Epub 2022 Apr 4. Microbiol Spectr. 2022. PMID: 35377226 Free PMC article.
-
Mechanisms of azole resistance in clinical isolates of Candida glabrata collected during a hospital survey of antifungal resistance.Antimicrob Agents Chemother. 2005 Feb;49(2):668-79. doi: 10.1128/AAC.49.2.668-679.2005. Antimicrob Agents Chemother. 2005. PMID: 15673750 Free PMC article.
-
A new 1-nitro-9-aminoacridine derivative targeting yeast topoisomerase II able to overcome fluconazole-resistance.Bioorg Med Chem Lett. 2021 Mar 1;35:127815. doi: 10.1016/j.bmcl.2021.127815. Epub 2021 Jan 21. Bioorg Med Chem Lett. 2021. PMID: 33486051
-
Candida and candidaemia. Susceptibility and epidemiology.Dan Med J. 2013 Nov;60(11):B4698. Dan Med J. 2013. PMID: 24192246 Review.
-
Resistance in human pathogenic yeasts and filamentous fungi: prevalence, underlying molecular mechanisms and link to the use of antifungals in humans and the environment.Dan Med J. 2016 Oct;63(10):B5288. Dan Med J. 2016. PMID: 27697142 Review.
Cited by
-
Anticancer drugs targeting topoisomerase II for antifungal treatment.Sci Rep. 2025 Mar 18;15(1):9311. doi: 10.1038/s41598-025-93863-z. Sci Rep. 2025. PMID: 40102495 Free PMC article.
References
-
- Staniszewska M, et al. Pathogenesis and treatment of infections. Postep. Mikrobiol. 2014;53:229–240.
-
- Skrodenienė E, Dambrauskienė A, Vitkauskienė A. Susceptibility of yeasts to antifungal agents in Kaunas University of Medicine Hospital. Med. 2006;42:294–299. - PubMed
-
- Biliński P, Seferyńska I, Warzocha K. Diagnosis and treatment of systemic fungal infections in oncohematology. Onkol. w Prakt. Klin. 2008;4:15–24.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
