

This is a preprint.
Understanding species-specific and conserved RNA-protein interactions in vivo and in vitro
- PMID: 38352439
- PMCID: PMC10862761
- DOI: 10.1101/2024.01.29.577729
Understanding species-specific and conserved RNA-protein interactions in vivo and in vitro
Update in
-
Understanding species-specific and conserved RNA-protein interactions in vivo and in vitro.Nat Commun. 2024 Sep 27;15(1):8400. doi: 10.1038/s41467-024-52231-7. Nat Commun. 2024. PMID: 39333159 Free PMC article.
Abstract
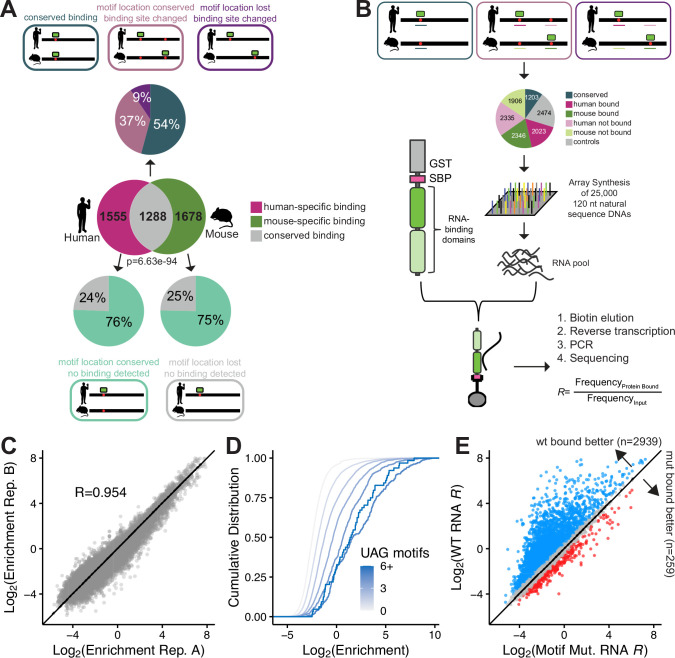
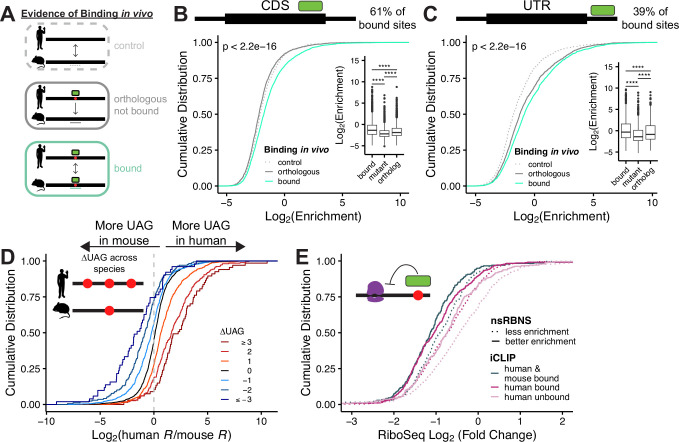
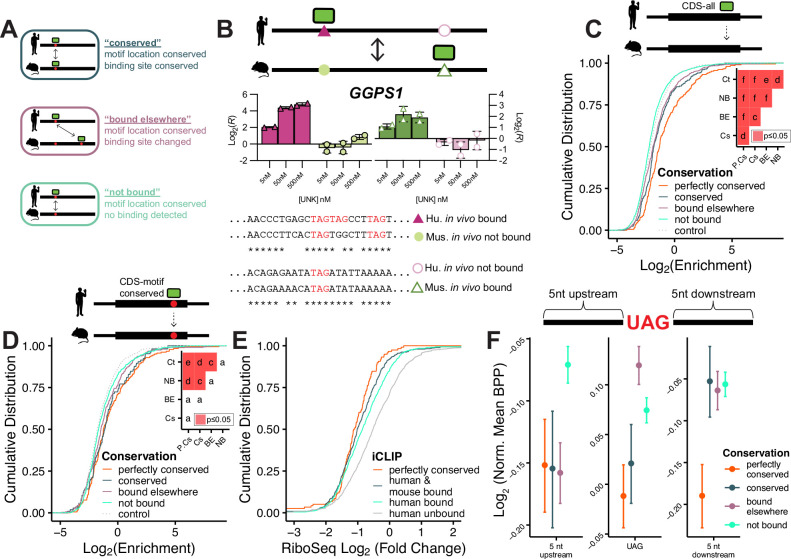
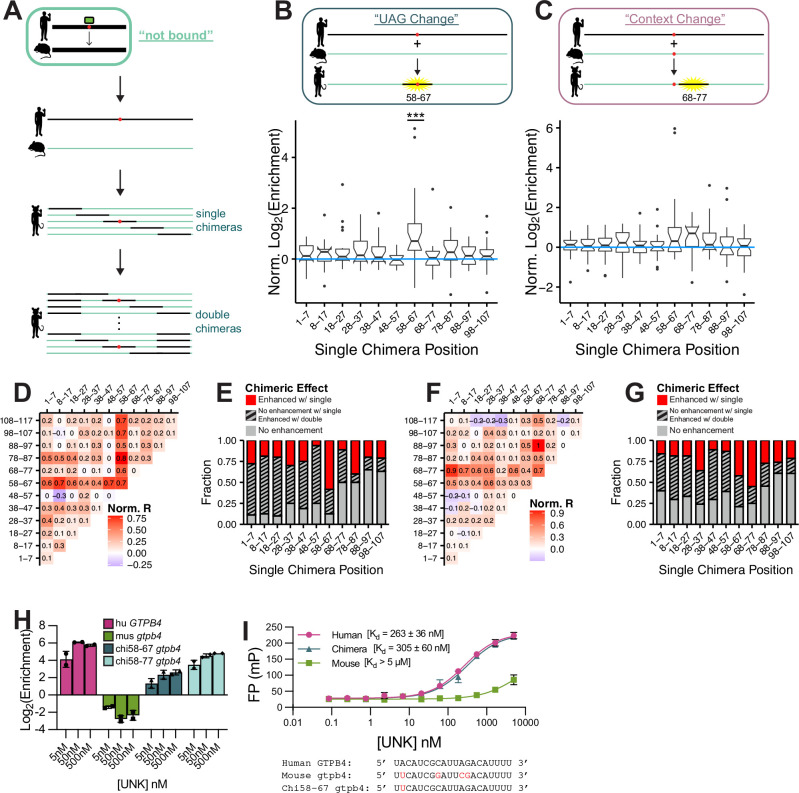
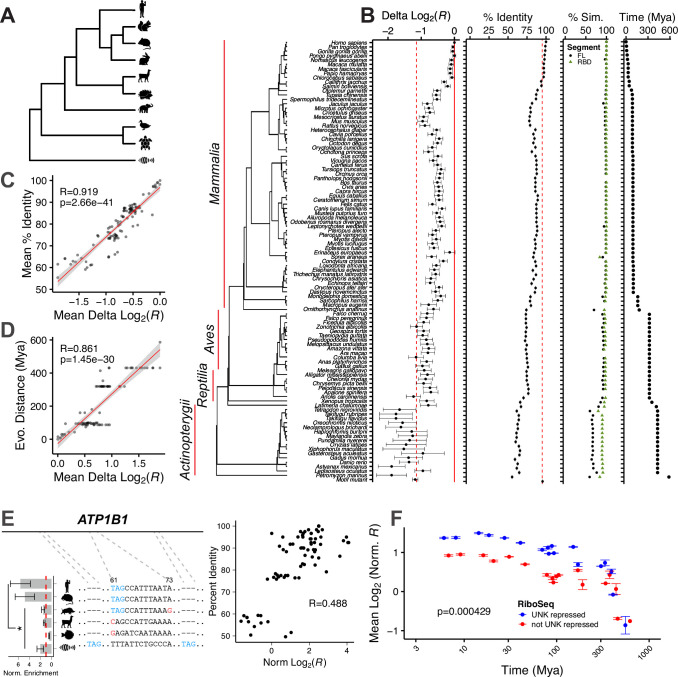
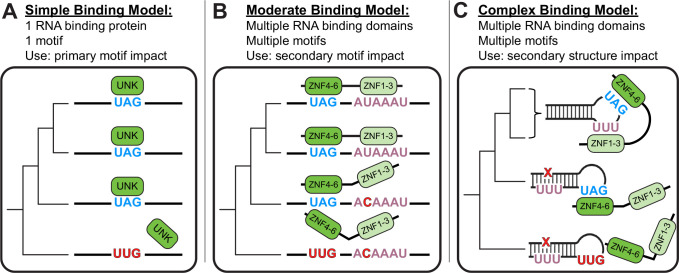
While evolution is often considered from a DNA- and protein-centric view, RNA-based regulation can also impact gene expression and protein sequences. Here we examined interspecies differences in RNA-protein interactions using the conserved neuronal RNA binding protein, Unkempt (UNK) as model. We find that roughly half of mRNAs bound in human are also bound in mouse. Unexpectedly, even when transcript-level binding was conserved across species differential motif usage was prevalent. To understand the biochemical basis of UNK-RNA interactions, we reconstituted the human and mouse UNK-RNA interactomes using a high-throughput biochemical assay. We uncover detailed features driving binding, show that in vivo patterns are captured in vitro, find that highly conserved sites are the strongest bound, and associate binding strength with downstream regulation. Furthermore, subtle sequence differences surrounding motifs are key determinants of species-specific binding. We highlight the complex features driving protein-RNA interactions and how these evolve to confer species-specific regulation.
Conflict of interest statement
Conflict of Interests The authors declare no conflicts.
Figures
References
-
- King M.-C. & Wilson A. C. Evolution at two levels in humans and chimpanzees: Their macromolecules are so alike that regulatory mutations may account for their biological differences. Science 188, 107–116 (1975). - PubMed
-
- Britten R. J. & Davidson E. H. Gene Regulation for Higher Cells: A Theory: New facts regarding the organization of the genome provide clues to the nature of gene regulation. Science 165, 349–357 (1969). - PubMed
-
- Britten R. J. & Davidson E. H. Repetitive and non-repetitive DNA sequences and a speculation on the origins of evolutionary novelty. The Q. review biology 46, 111–138 (1971). - PubMed
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources