

This is a preprint.
Spike transmission failures in axons from mouse cortical pyramidal neurons in vivo
- PMID: 38352485
- PMCID: PMC10862735
- DOI: 10.1101/2024.01.29.577733
Spike transmission failures in axons from mouse cortical pyramidal neurons in vivo
Update in
-
Spike transmission failures in axons from cortical neurons in vivo.iScience. 2024 Sep 5;27(10):110884. doi: 10.1016/j.isci.2024.110884. eCollection 2024 Oct 18. iScience. 2024. PMID: 39346673 Free PMC article.
Abstract
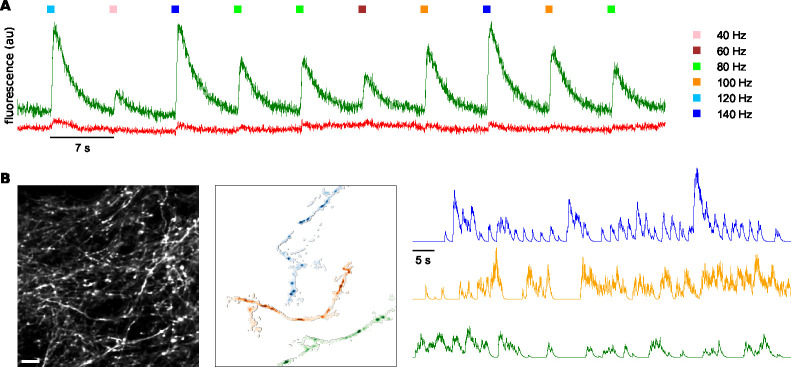
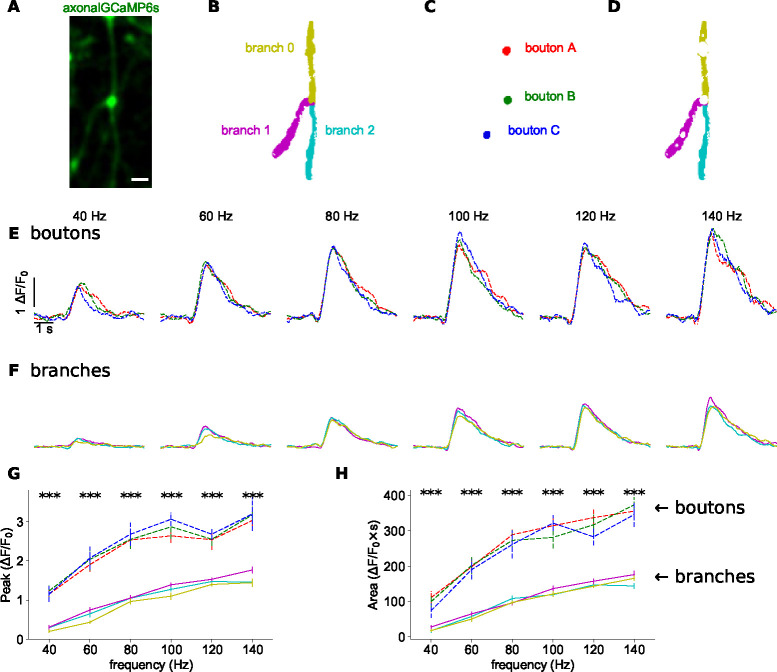
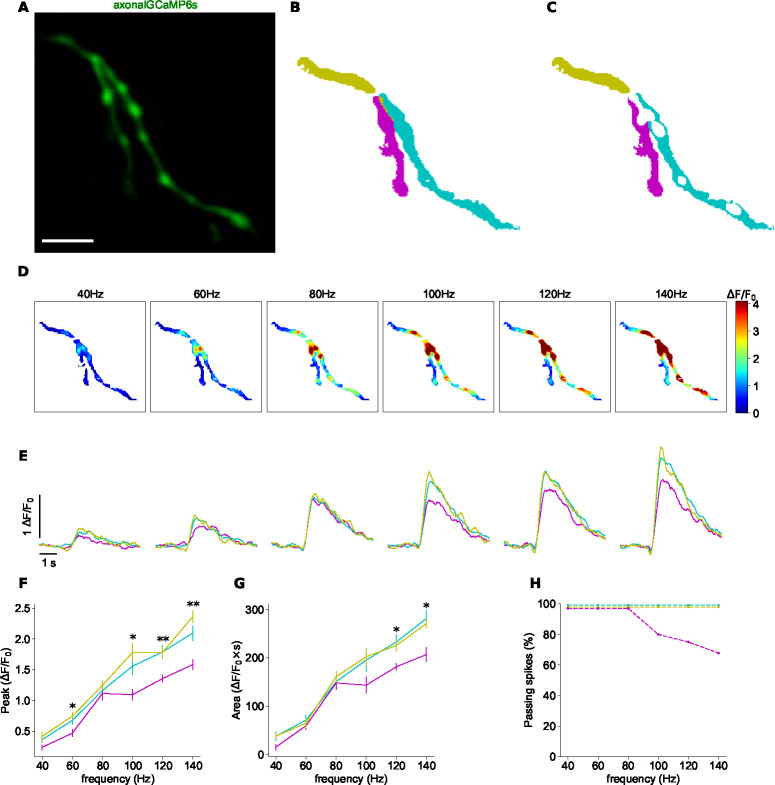
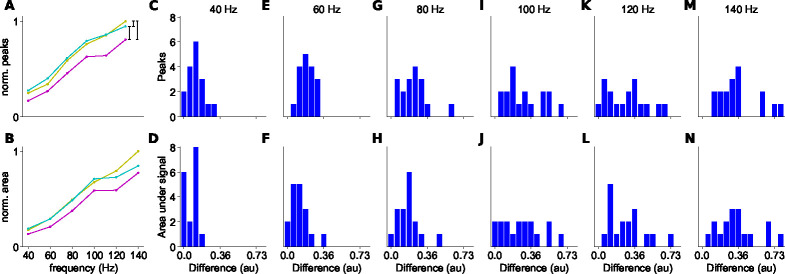
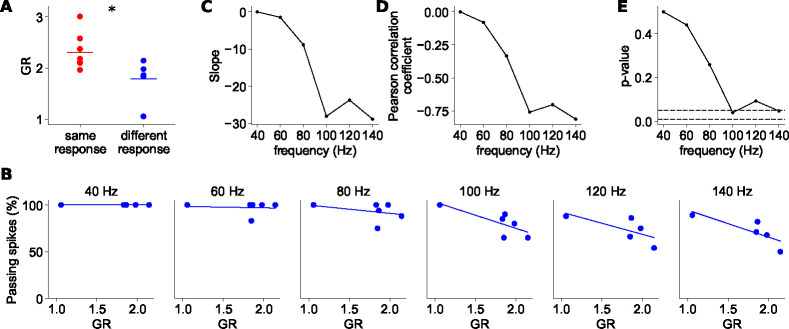
The propagation of action potentials along axons is traditionally considered to be reliable, as a consequence of the high safety factor of action potential propagation. However, numerical simulations have suggested that, at high frequencies, spikes could fail to invade distal axonal branches. Given the complex morphologies of axonal trees, with extensive branching and long-distance projections, spike propagation failures could be functionally important. To explore this experimentally in vivo, we used an axonal-targeted calcium indicator to image action potentials at axonal terminal branches in superficial layers from mouse somatosensory cortical pyramidal neurons. We activated axons with an extracellular electrode, varying stimulation frequencies, and computationally extracted axonal morphologies and associated calcium responses. We find that axonal boutons have higher calcium accumulations than their parent axons, as was reported in vitro. But, contrary to previous in vitro results, our data reveal spike failures in a significant subset of branches, as a function of branching geometry and spike frequency. The filtering is correlated with the geometric ratio of the branch diameters, as expected by cable theory. These findings suggest that axonal morphologies contribute to signal processing in the cortex.
Figures

References
-
- Chéreau R, Saraceno GE, Angibaud J, Cattaert D, Nägerl UV. Superresolution imaging reveals activity-dependent plasticity of axon morphology linked to changes in action potential conduction velocity. 2017. Proceedings of the National Academy of Sciences of the United States of America; 114(6):1401–1406 doi: 10.1073/pnas.1607541114. - DOI - PMC - PubMed
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources