

The B7:CD28 family and friends: Unraveling coinhibitory interactions
- PMID: 38354702
- PMCID: PMC10889489
- DOI: 10.1016/j.immuni.2024.01.013
The B7:CD28 family and friends: Unraveling coinhibitory interactions
Abstract
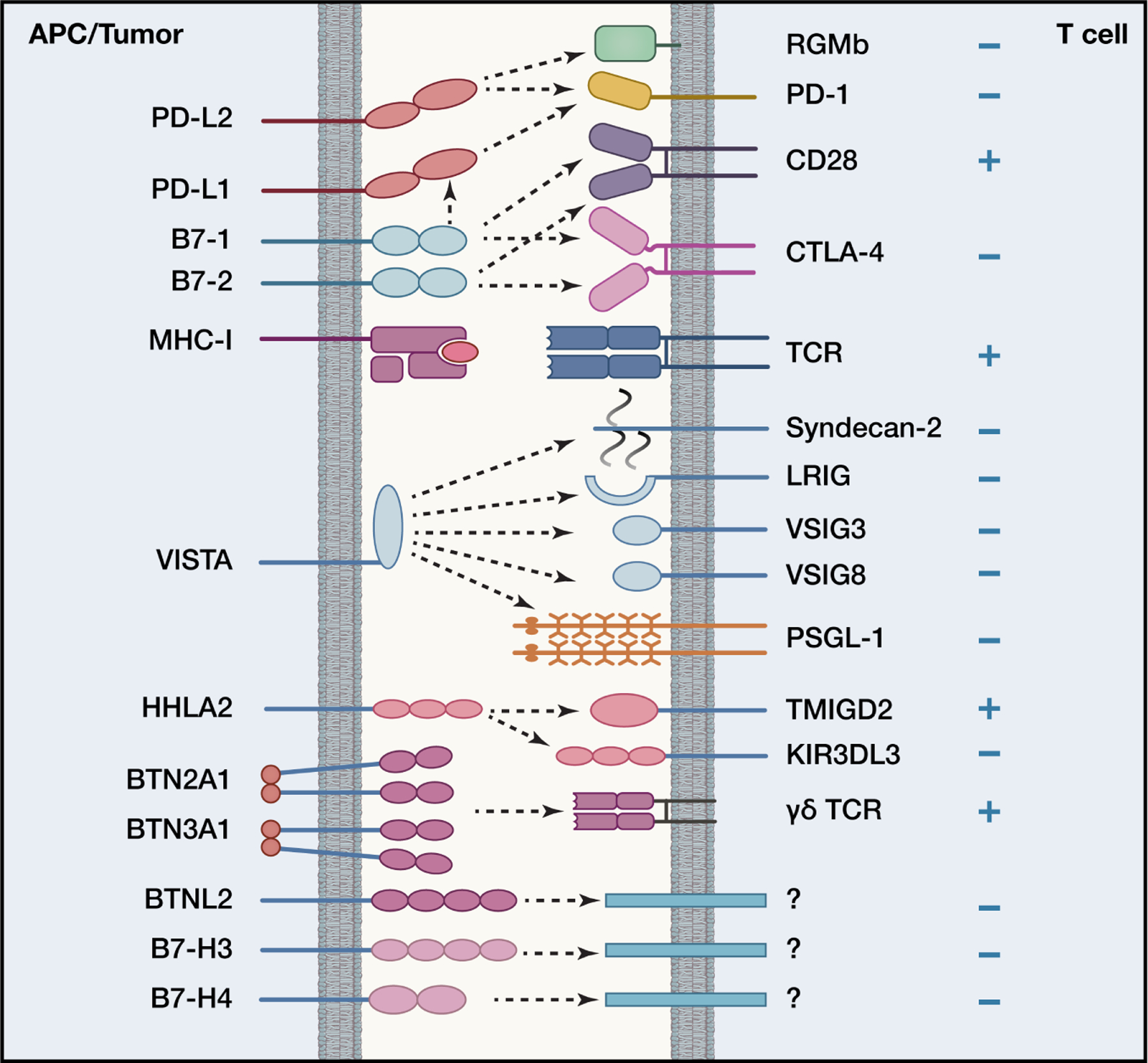
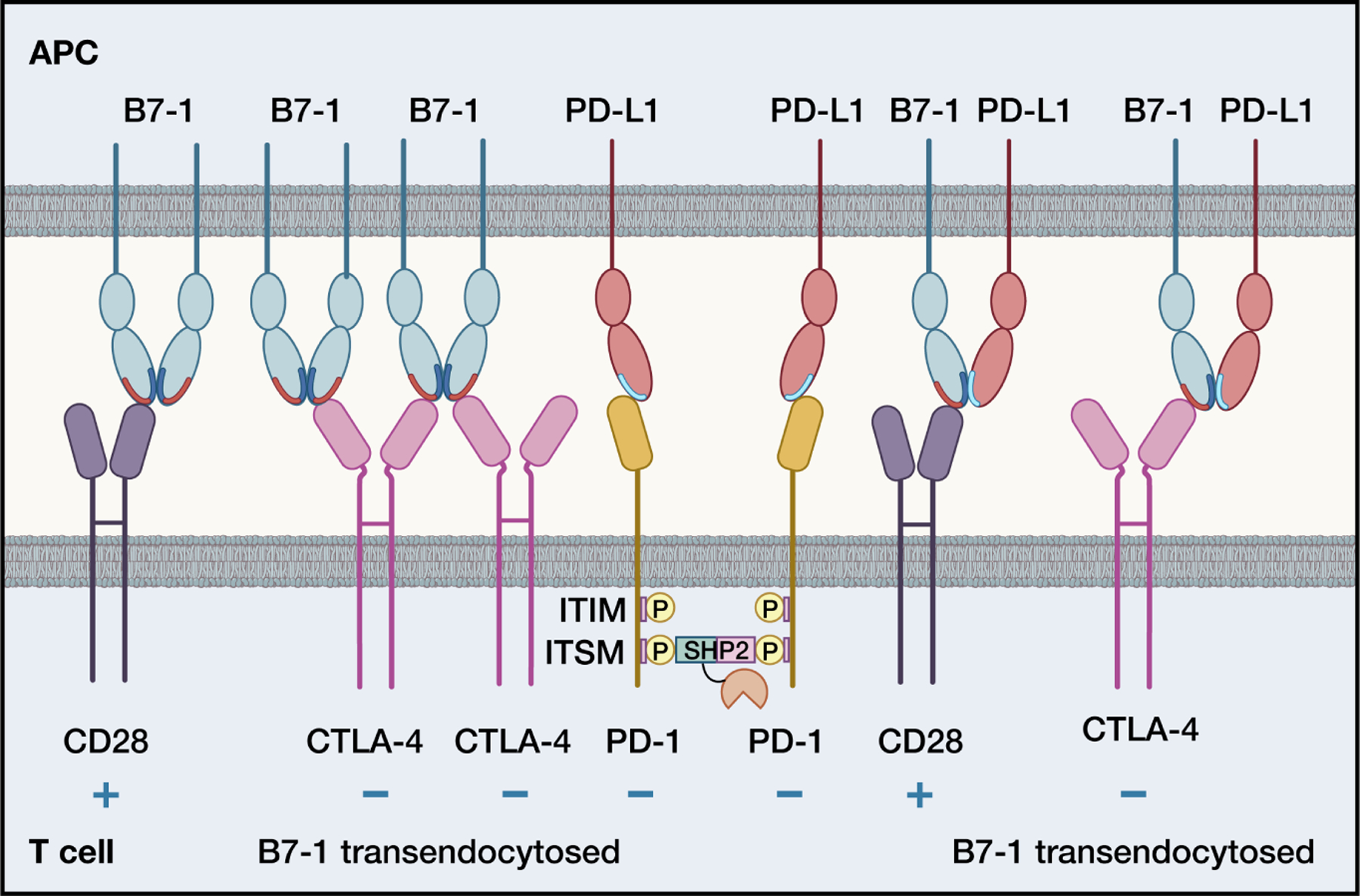
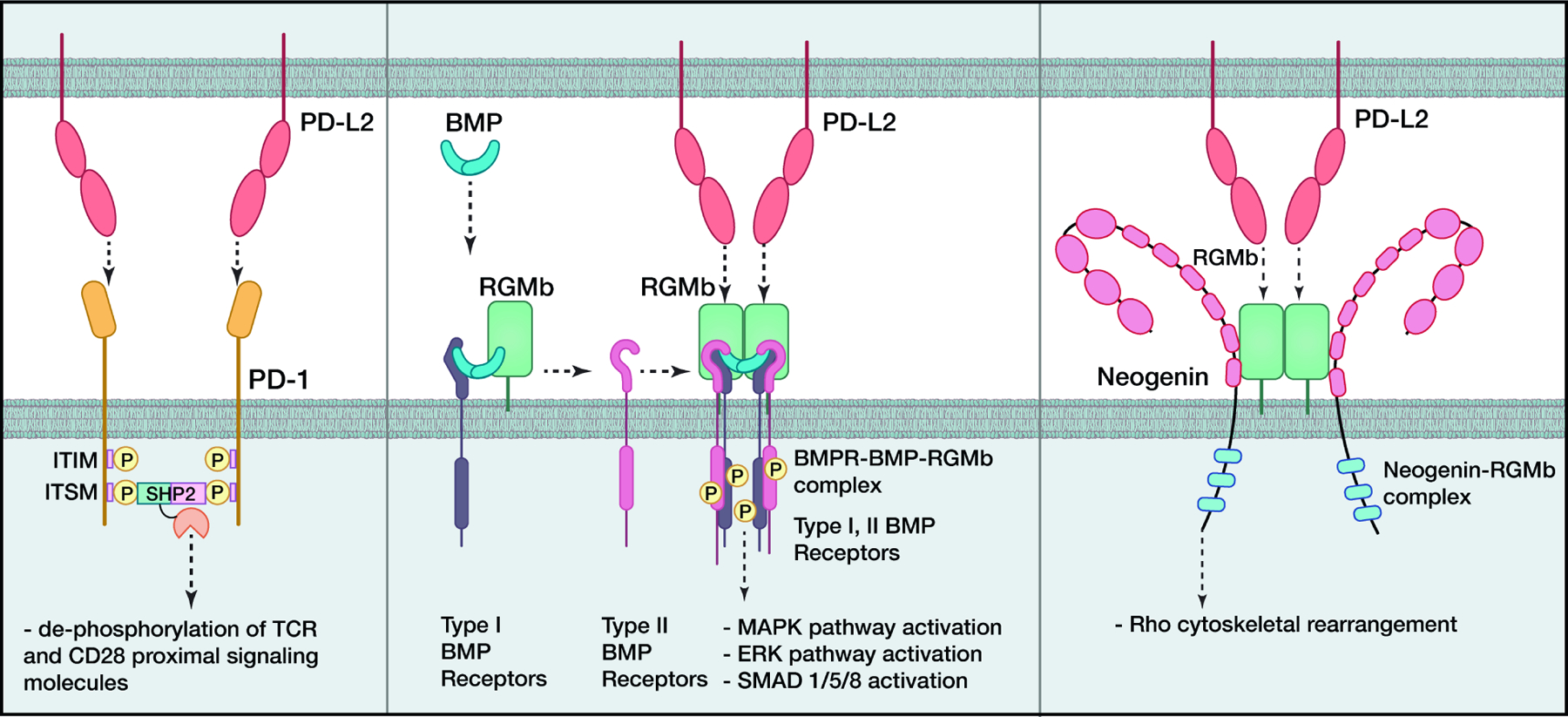
Immune responses must be tightly regulated to ensure both optimal protective immunity and tolerance. Costimulatory pathways within the B7:CD28 family provide essential signals for optimal T cell activation and clonal expansion. They provide crucial inhibitory signals that maintain immune homeostasis, control resolution of inflammation, regulate host defense, and promote tolerance to prevent autoimmunity. Tumors and chronic pathogens can exploit these pathways to evade eradication by the immune system. Advances in understanding B7:CD28 pathways have ushered in a new era of immunotherapy with effective drugs to treat cancer, autoimmune diseases, infectious diseases, and transplant rejection. Here, we discuss current understanding of the mechanisms underlying the coinhibitory functions of CTLA-4, PD-1, PD-L1:B7-1 and PD-L2:RGMb interactions and less studied B7 family members, including HHLA2, VISTA, BTNL2, and BTN3A1, as well as their overlapping and unique roles in regulating immune responses, and the therapeutic potential of these insights.
Copyright © 2024 Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests A.H.S. has patents/pending royalties on the PD-1 pathway from Roche and Novartis. A.H.S. is on advisory boards for Elpiscience, Bicara, Monopteros, Fibrogen, Alixia, IOME, Corner Therapeutics, Bioentre, GlaxoSmithKline, Amgen, and Janssen. She also is on scientific advisory boards for the Massachusetts General Cancer Center, Program in Cellular and Molecular Medicine at Boston Children’s Hospital, the Human Oncology and Pathogenesis Program at Memorial Sloan Kettering Cancer Center, the Johns Hopkins Bloomberg Kimmel Institute for Cancer Immunotherapy, and the Gladstone Institute and is an academic editor for the Journal of Experimental Medicine. A.H.S. has funding from Quark/Iome; AbbVie; Calico; and Taiwan Bio. G.J.F. has patents/pending royalties on the PD-L1/PD-1 pathway from Roche, Merck MSD, Bristol-Myers-Squibb, Merck KGA, Boehringer-Ingelheim, AstraZeneca, Dako, Leica, Mayo Clinic, Eli Lilly, and Novartis. G.J.F. has patents or patent applications on the use of PD-L1, PD-L2, PD-1, RGMb, HHLA2, KIR3DL3, and BTNL2 in cancer immunotherapy. G.J.F. has served on advisory boards for iTeos, NextPoint, IgM, GV20, IOME, Bioentre, Santa Ana Bio, Simcere of America, and Geode. G.J.F. has equity in Nextpoint, Triursus, Xios, iTeos, IgM, Trillium, Invaria, GV20, Bioentre, and Geode.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Research Materials
