

SLC25A51 decouples the mitochondrial NAD+/NADH ratio to control proliferation of AML cells
- PMID: 38354740
- PMCID: PMC10990793
- DOI: 10.1016/j.cmet.2024.01.013
SLC25A51 decouples the mitochondrial NAD+/NADH ratio to control proliferation of AML cells
Abstract
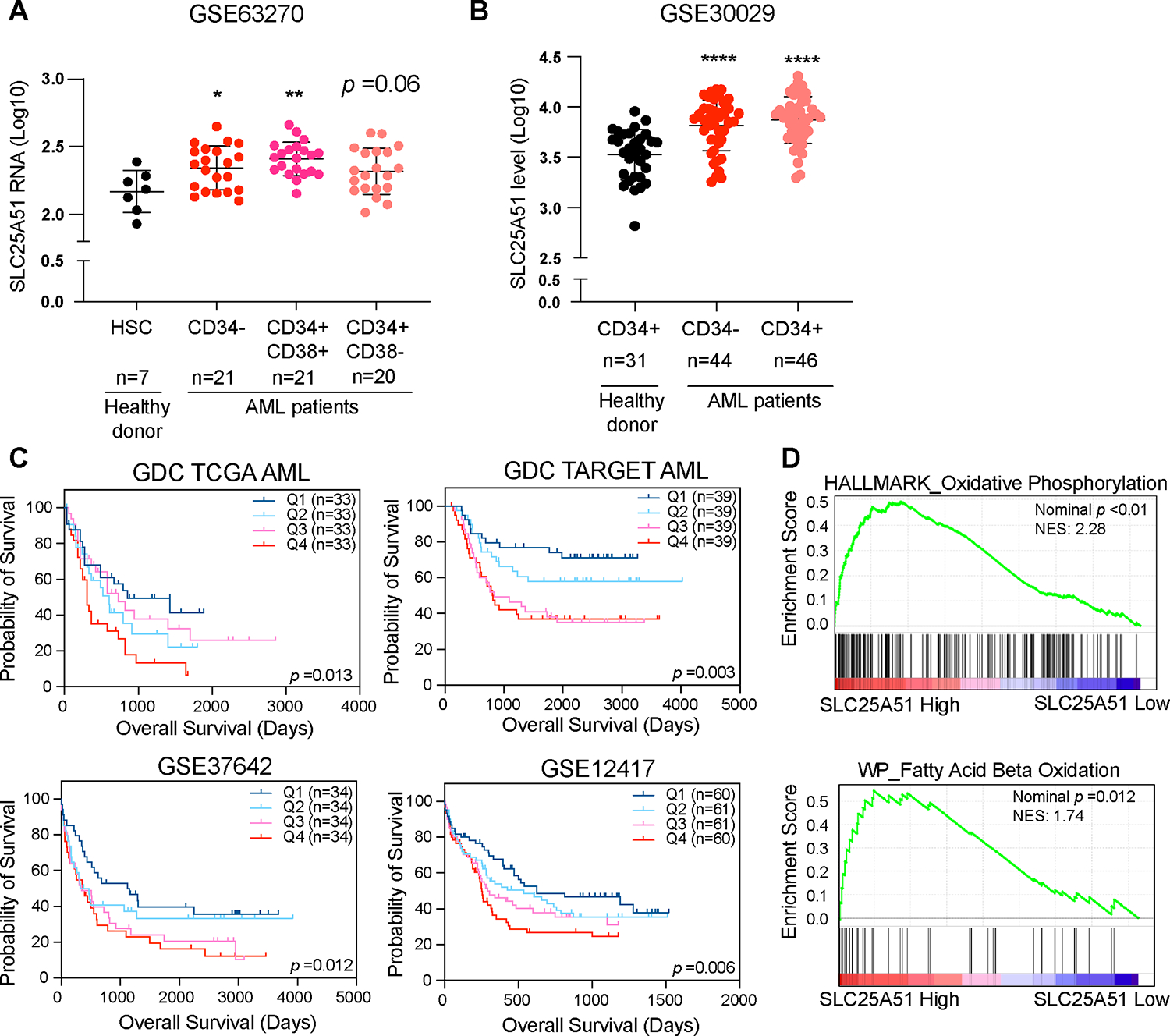
SLC25A51 selectively imports oxidized NAD+ into the mitochondrial matrix and is required for sustaining cell respiration. We observed elevated expression of SLC25A51 that correlated with poorer outcomes in patients with acute myeloid leukemia (AML), and we sought to determine the role SLC25A51 may serve in this disease. We found that lowering SLC25A51 levels led to increased apoptosis and prolonged survival in orthotopic xenograft models. Metabolic flux analyses indicated that depletion of SLC25A51 shunted flux away from mitochondrial oxidative pathways, notably without increased glycolytic flux. Depletion of SLC25A51 combined with 5-azacytidine treatment limits expansion of AML cells in vivo. Together, the data indicate that AML cells upregulate SLC25A51 to decouple mitochondrial NAD+/NADH for a proliferative advantage by supporting oxidative reactions from a variety of fuels. Thus, SLC25A51 represents a critical regulator that can be exploited by cancer cells and may be a vulnerability for refractory AML.
Keywords: AML; MCART1; SLC25A51; glutamine utilization; oxidative mitochondria; tumor metabolism.
Copyright © 2024 Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests The authors declare no competing interests.
Figures





References
-
- Pachnis P, Wu Z, Faubert B, Tasdogan A, Gu W, Shelton S, Solmonson A, Rao AD, Kaushik AK, Rogers TJ, et al. (2022). In vivo isotope tracing reveals a requirement for the electron transport chain in glucose and glutamine metabolism by tumors. Sci. Adv. 8, eabn9550. 10.1126/sciadv.abn9550. - DOI - PMC - PubMed
-
- Garcia-Bermudez J, Baudrier L, La K, Zhu XG, Fidelin J, Sviderskiy VO, Papagiannakopoulos T, Molina H, Snuderl M, Lewis CA, et al. (2018). Aspartate is a limiting metabolite for cancer cell proliferation under hypoxia and in tumours. Nat Cell Biol 20, 775–781. 10.1038/s41556-018-0118-z. - DOI - PMC - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
